"How to reach LEVEL 5 in autonomous vehicles, robots, cars etc."
US 6172941 (filing date: Dec. 16, 1999,
granted 2001) see Google Patents
EP 1146406A1 (filing
date: Dec. 03, 1999)
Inventor & Author: Erich Bieramperl, 4040 Linz, Austria - EU
Elapse Time Quantizing
Autoadaption-Theorem
Algorithm of Life
The Neuronal Code
The Meaning of the Tetragrammaton JHWH / YHVH
A
Step to a New Universal Theory?
A method to generate recognition, auto-adaptation and self-organization in autonomous mechanisms and organisms. A number of sensing elements generate analog signals whose amplitudes are classified into different classes of perception intensity. The currently occurring elapse times between phase transitions are recorded and compared with prior recorded elapse times in order to find covariant time sequences and patterns. A motion actuating system can be coupled to the assembly, which is controlled by pulse sequences that have been modulated in accordance with the covariant time sequences. In this way the mechanism or organism in motion is prompted to emulate the found covariant time sequences, while being able to recognize its own motion course and adapting itself to changes of environment.
Background
This invention describes a method for generating processes
that facilitate the self-organization ofautonomous systems. It can
be applied to mechanistic fields as well as to molecular/biological
systems. By means of the invention described herein, it is possible
for a system in motion to recognize external events in a subjective
way through self-observation; to identify the surrounding physical
conditions in real time; to reproduce and to optimize the system's
own motions; and to enable a redundancy-poor process that leads to
self-organization.
Robot systems of the usual static type are
mainly based on deterministic path dependent regulating processes.
The digital outputs and values that control the robot's position are
stored in the memory of a central computer. Many degrees of freedom
can be created by a suitable arrangement of coordinating devices.
Position detectors can be devices such as tacho-generators, encoders,
or barcode rulers scanned by optical sensors that provide path
dependent increment pulses. The activation mostly takes place by
means of stepper motors.
It is also well-known that additional
adaptive regulating processes based on discrete time data are used
in path dependent program control units. These data are produced by
means of the SHANNON-quantization method, utilizing
analog-to-digital converters to sample the amplitudes of sensors and
transducers. They serve to identify the system's actual value (i.e.
its current state). Continued comparison of reference values and
actual values are necessary for correction and adjustment of the
regulating process. Newly calculated parameters are then stored in
the memory. This kind of adaptive regulation is necessary, for
example, in order to eliminate a handling robot's deviations from a
pre-programmed course that are caused by variable load conditions.
If a vehicle that is robot-controlled in this way were to be placed
into an autonomous state, it would generally be impossible to
determine its exact position reference (i.e. coordinates) by means
of tachogenerators or encoders. For this reason controlling values
(or commands) cannot be issued by a computer - or preprogrammed into
a computer - in an accurate manner. This is true not only for
robot-controlled automobiles, gliding vehicles, hovercraft or
aircraft, but also for rail-borne vehicles for which the distance
dependent incremental pulses are often inaccurate and therefore not
reproducible. This is usually caused by an uneven surface or worn or
slipping wheels. Explorer robots, which are used to locate objects
or to rescue human beings from highly inaccessible or dangerous
locations, must therefore be controlled manually, or with computer
supported remote control units. A video communication system is
necessary for such cases in order to be able to monitor the motion
of the robot. However, in many applications of robotics, this is
inadequate. A robot-controlled automobile, for example, should be
capable of avoiding dangerous situations in real time, as well as
being capable of adapting its speed to suit the environment, without
any human intervention. In such cases, it is necessary for the
on-board computer to recognize the situation at hand, then calculate
automatically the next steps to be carried out.
In this way the
robot-controlled vehicle ought to have a certain capability for
self-organization. This is also true for other robot-controlled
systems.
With regards to autonomous robot systems, techniques
already exist to scan the surroundings by means of sensors and to
analyze the digital sensor data that were acquired using the
above-mentioned discrete time quantization method (see Fig. 1); and
there already exist statistical calculation methods and algorithms
that generate suitable regulating parameters. Statistical methods
for handling such regulating systems were described in 1949 by
Norbert WIENER. According to the SHANNON theorem, the scanning of
the external environment must be done with at least double the
frequency of the signal amplitude bandwidth. In this way the
information content remains adequate. In order to be able to
identify the robot's own motions, very high sampling rates are
necessary. This amplitude quantization method currently in
widespread use requires the correlation of particular measurement
data to particular points in time (Ts) that are predetermined using
the program counter. For this reason this should be understood as a
deterministic method. However, practical experience has shown that
even ultrahigh-speed processors and the highest sampling rates
cannot provide sufficient efficiency. The number of redundant data
and the amount of computing operations increase drastically when a
moving sensor-controlled vehicle meets new obstacles or enters new
surroundings at variable speed. Indeed, C. SHANNON's quantization
method does not allow recognition of an analogue signal amplitude in
real time, especially if there are changing physical conditions or
variable motions for which the acquisition of additional information
regarding the instantaneous velocity is necessary. This is also true
if laser detectors or supersonic sensors are used, for which mainly
distance data are acquired and processed.
Therefore, although
this quantization method is suitable for analyzing the trace of a
motion and for representing this motion on a monitor (see Pat.
AT
397 869), it is hardly adequate for recognizing the robot's own
motion, or for reproducing it in a self-adaptive way.Some autonomous
mobile robot systems operate with CCD sensors and OCR software (i.e.
utilising image processing). These deduce contours or objects from
color contrast and brightness differentials, which are interpreted
by the computer as artificial horizons or orientation marks.
Examples of this technology are computer-supported guidance systems
and steering systems that allow vehicles to be guided automatically
by centre lines, side planks, street edges and so on. CCD sensors -
when one observes how they operate - are analog storage devices that
function like well-known bucket brigade devices. Tightly packed
capacitors placed on a MOS silicon semiconductor chip are charged by
the photoelectric effect to a certain electrical potential. Each
charge packet represents an individual picture element, termed
"pixel"; and the charge of each pixel is a record of how bright that
part of the image is. By supplying a pulse frequency, the charges
are shifted from pixel to pixel across the CCD, where they appear at
the edge output as serial analog video signals. In order to process
them in a computer, they must be converted into digital quantities.
This requires a large number of redundant data and calculations;
this is why digital recording of longer image sequences necessitates
an extremely large high speed memory. Recognizing isomorphous
sequences in repetitive motions is only possible with large memory
and time expenditure, which is why robotic systems based on CCD
sensors cannot adequately reproduce their own motion course in a
self-adaptive way. With each repetition of the same motion along the
same route, all regulating parameters must be calculated by means of
picture analysis anew. If environment conditions change through fog,
darkness or snowfall, such systems are overburdened.
Pat. AT 400 028
describes a system for the adaptive regulation of a motor driven
vehicle, in which certain landmarks or signal sources are provided
along the vehicle's route in order to serve as bearing markers that
allow the robot to keep to a schedule. Positions determined by GPS
data can also serve this purpose. When the system passes these
sources, the sensor coupled on board computer acquires the elapsed
times for all covered route segments by means described in
Pat. U.S.
4,245.334, which details the manner of time quantization by first
and second sensor signals. The data acquired in this way serve as a
reference base for the computation of regulating parameters that
control the drive cycles and brake cycles of the vehicle when a
motion repetition happens. The system works with low data
redundancy, corrects itself in a self-adaptive manner, and is
capable of reproducing an electronic route schedule precisely. It is
suitable, for example, for ensuring railway networks keep to
schedule. However, in the system detailed in the above-mentioned
patent, it is not possible to identify external objects and
surroundings.
It is an object of the present invention to provide
an extensive method for the creation of autonomous self-organizing
robot systems or organisms, which enables them to identify external
signals, objects, events, physical conditions or surroundings in
real time by observing from their own subjective view. They will be
able to recognize their own motion patterns and to reproduce and
optimize their behavior in a self-adaptive way. Another object of
this invention is the preparation of an autonomous training robot
for use in sports, that is capable of identifying, reproducing and
optimizing a motion process (e.g. that has been trialed beforehand
by an athlet) as well as: determining the ideal track and speed
courses automatically; keeping to route schedules; representing its
own motion, speeds, lap times, intermediate times and start to
finish times on a monitor; and which is capable of acoustic or
optical output of the acquired data.
Summary Of The Invention
The requirements outlined in the previous paragraph
are solved generically by attaching analog sensors or receptors onto
the moving system (for example, a robot system) which scan
surrounding signal sources whose amplitudes are subdivided by
defining a number of threshold values. This creates perception
zones. The elapsed times of all phase transitions in all zones are
measured by means of analog or digital STQ quantization, and the
frequency of the time pulses is modulated automatically, depending
on the relative instantaneous speed which is determined by the phase
displacement of equi-valent sensors.Therefore the counted time
pulses correlate approximately with the length-values d(nnn). With
this method, the scanning of signal amplitudes is not a
deterministic process: it is not carried out at predetermined times
with predetermined time pulses. The recording, processing and
analysis of the elapsed times takes place according to probabilistic
principles. As a result, a physically significant phenomenon arises:
the parameters describing the external surroundings are not
objectively measured by the system, but are subjectively sensed as
temporal sequences. The system itself functions as observer of the
process. In the technical literature - in the context of
deterministic timing - elapse times are also
called "signal
running times" or "time intervals ". According to the present
invention, the so-called
STQ elapse times in a signal-recognition
process are quantized with every transition of a phase amplitude
through a threshold value (which is effected by starting and
stopping a number of timers). This produces a stream of time data.
Every time elapsed between phase transitions in the "equal zone", as
well as the time elapsed between transitions through a low threshold
value then a higher threshold value (and vice versa), can be
recorded.
STQ(v) = sensitivity/ time quantum of
velocity = Tv1,2,3...
This is the elapsed
time determined by the signal amplitude that occurs when a first
sensor (or receptor). S2 and an equivalent second sensor (or
receptor) S1 moves along a corresponding external signal source Q,
measured from the rising signal edge at the phase transition iTv1.1
of the first sensor signal to the rising signal edge at the phase
transition iTw1.1 of the second sensor signal; and likewise from
iTv2.1 to iTw2.1, from iTv3.1 to iTw3.1. (These transitions
correspond to equivalent threshold values P1,2,3...) STQ(v) times
can also be measured from falling signal edges. They serve as
parameters for the immediate relative velocity (vm) of the system in
motion.
STQ(d) =
sensitivity/time quantum of differentiation = Td1,2,3...
This is the elapsed time determined by the signal amplitude of a
sensor (or receptor) S within range of a corresponding external
signal source (Q1,2,3..), measured from the rising signal edge at
the phase transition iTw1 of a rising amplitude trace to the rising
signal edge at the next higher phase transition iTw2, and from the
rising edge at iTw2 to the rising edge at iTw3, from the rising edge
at iTw3 to the rising edge at iTw4, and so on; or, equivalently,
from successive falling edges when amplitude traces are falling.
(These transitions correspond to the equivalent threshold values
P1,2,3,4..) STQ(d) elapse times are differentiation parameters for
the slope of signal amplitudes (and consequently for their
frequency); furthermore they serve as a plausibility check and
verification of other corresponding STQ data. With this measurement,
the relative motion between sensor and signal source is not taken
into account. In the case of no relative motion between sensors and
sources, changes in the source field are detectable and recognizable
by recording STQ(i) and/or STQ(d) data. If the source field is
invariant, a recognition is only possible if STQ(i) or STQ(v)- data
are derived from variable threshold values (focusing). If there is
absolute physical invariance, no STQ-quantum can be acquired, and
recognition is impossible. STQ(v)-data are recorded in order to
recognize the spatial surroundings under relative motion, and/or to
identify relative motion processes so as to be able to recognize
the self-motion (or components of this motion); as well as to
reproduce any motion in a self-adaptive manner.
If the method
presently being described is applied in a mechanistic area, the
above-mentioned perception area zones may normally be set by a
number of electronic threshold value detectors with pre-definable
threshold levels, and the STQ(i) and STQ(d) elapse time data are
acquired by programmable digital timers. The elapse timing process
is actuated at an iT phase transition as well as halted at an iT
phase transition. Then the time data are stored in memory.
Moreover, these STQ(v) elapse times are recorded by means of
electronic integrators, in which the charge times of the capacitors
determine those potentials that are applied as analog STQ(v) data to
voltage/frequency converters, in order to modulate the digital time
pulse frequencies for the adaptive measurement of STQ(i) and STQ(d)
data, in a manner which is a function of the relative speed vm.
In non-mechanistic implementations of the method presently being
described, it is intended that the so-called perception area zones,
as well as the threshold value detectors and the previously
described STQ-quantization processes, are not formed in the same
manner as in electronic analog/digital circuits, but in a manner
akin to molecular/biological structures. In other general
implementations, it is intended that those time stream patterns that
consist of currently recorded STQ data be continuously compared with
prior recorded time stream patterns by means of real time analysis,
in order to identify external events or changes in physical
surroundings with a minimum of redundancy, as well as to recognize
these in real time.
In yet another possible general
implementation, it is intended that autonomously moving systems,
that are equipped with sensors and facilities capable of the kind of
time stream pattern recognition mentioned above, have propulsion,
steering and brake mechanisms that are regulated in such a manner,
that the autonomously moving system (in particular, a mobile robot
system) is capable of reproducing prior recorded STQ time stream
patterns in a self-adaptive way. When repeating this movement, a
processor deletes unstable or insufficiently co-ordinated time
stream data from memory, while assigning only those time stream data
as instruction, which allows reproduction of the motion along the
same routes in an optimal co-ordinated manner.
In addition, it is
intended that the time base frequency for the above mentioned STQ
elapse timing is increased or decreased in order to scale the time
sequences proportionally, whereby the velocity of all movements is
proportionally scaled too.
Short Description Of The Figures
Fig. 1 shows a diagram of SHANNON's
deterministic method of discrete time quantization of signal
amplitude traces.
Figs. 2a-c are graphic
diagrams of the quantization of signal amplitude traces by means of
acquisition of STQ(v), STQ(i) and STQ(d) elapse times, according to
the herein described non-deterministic method.
Figs.
3a-c illustrate this non-deterministic quantization method
in connection with serial transfer of acquired STQ(d)- elapse times,
as well as time pulse frequency modulation of simultaneously
acquired parameters of the immediate relative speed (vm).
Figs. 3d-g illustrate, in accordance with the
presently described invention, a method to compare the currently
acquired STQ time data sequences with prior recorded STQ time data
sequences, in order to detect isomorphism of certain time stream
patterns.
Fig. 4a shows an action potential
AP
Fig. 4b shows vm dependent action
potentials which propagate from a sensory neuron (receptor) along a
neural membrane to the synapse where the covariance of STQ sequences
is analysed.
Fig. 4c shows a number of vm
dependent action potentials, which propagate from a group of
suitable receptors along collateral neural membranes to synapses, at
which the "temporal and spatial facilitation" of AP's is analysed
together with the covariances of these STQ sequences in order to
recognize a complex perception.
Fig. 4d
shows a postsynaptic neuron that produces potentials with inhibitory
effects.
Fig. 4e and Fig. 4f
show the general function of the synaptic transfer of
molecular/biologically recorded STQ information to other neurons or
neuronal branches.
Fig. 5 shows a
configuration where the described invented method has been applied
to generate an autonomous self-organizing mechanism, and where the
STQ time data are acquired by means of electronics.
Fig. 6a shows a configuration of a concrete embodiment of
the present method, where (as in Figs. 2a - 2c) the acquired STQ(v),
STQ(i) and STQ(d) time data are applied to the recognition of
certain spatial profiles, structures or objects when the system is
in motion at arbitrary speed.
Figs. 6b-e
illustrate several diagrams and schedules in accordance with the
particular embodiment in Fig. 6a, in which the sensory scanning and
recognition of certain profiles can occur under invariable or
variable speed course conditions.
Figs. 7a-d
show several configurations of sensors and sensor structures for the
recording of STQ(v) elapse times, which serve as parameters of the
immediate relative velocity vm.
Figs. 8a-f
illustrate a configuration, as well as the principles under which
another embodiment of the invention functions, in which the
acquisition of STQ time data (see Figs. 2a - 2c) is used to create
an autonomous self-adaptive and self-organizing training robot for
use in sport. This embodiment is capable of reproducing and
optimizing motion processes that have been pre-exercised by the
user. It is also capable of determining the ideal track and speed
courses automatically; of keeping distances and times; of
recognizing and warning in advance of dangerous impending
situations; and of representing on a monitor the self-motion, in
particular the speed, lap times, intermediate times, start to finish
times and other relevant data. In additional, this embodiment is
capable of displaying these acquired data in an optical or acoustic
manner.
Fig. 9 shows a schematic diagram of
the automatic focusing of certain perception zones or threshold
values, through which it is intended to improve and optimize the
recognition capability and the auto-covariant behaviour of the
system in motion. (This point is object of an additional patent
application).
Fig. 10 illustrates in a
general schematic view the production of time data streams by
amplitude transitions at certain sensory perception areas or
sensitivity zones (or threshold values, respectively) in autonomous
self-adaptive and self-organizing structures, organisms or
mechanistic robot systems, where a multiplicity of types of sensors
or receptors can exist.
Detailed Description Of The Invention
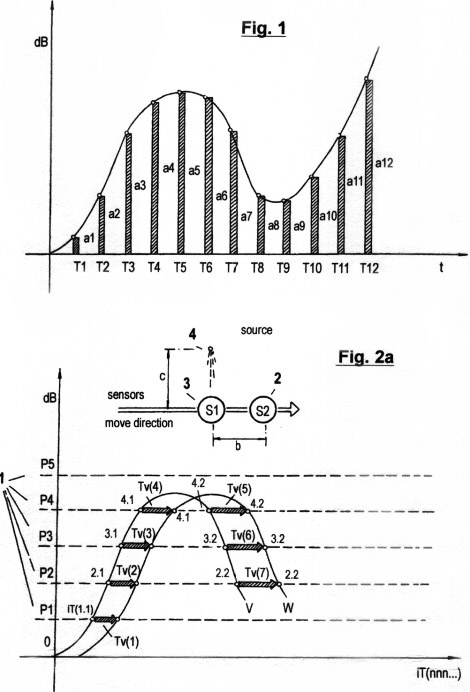
Fig. 1 shows a diagram of SHANNON's deterministic method of discrete time quantization of signal amplitude traces, which are digitized by analog/digital converters. In the usual technical language this method is called "sampling". This deterministic quantization method is characterized by quantized data (a1,a2,a3 ...an) which correlate to certain points in time (T1,T2,T3, ...Tn) that are predetermined from the program counter of a processor.
In present day robotics practice, this currently used deterministic method requires very fast processors, high sampling rates and highly redundant calculations for the processing and evaluation of data. If one wants to acquire sensor data from signal amplitudes of external sources for the purpose of getting information about the spatial surroundings of a system in which a sensor coupled processor is installed, SHANNON's method is incapable of generating suitable data for the immediate relative speed and temporal allocation, data which are necessary to optimize the coordination of the relative self-motion. A recognition of its own motion in real time therefore is not possible. For this reason, this currently used deterministic method is inadequate for the generation of highly effective autonomous robot systems.
Figs. 2a - c show three different graphs of direct "sensory quantization" of signal amplitude traces by means of the herein described invented method. In contrast to the quantization method shown in Fig. 1, in this method no vertical segments of amplitude traces are scanned; there are only elapse time measurements carried out in three different complementary ways. As is easy seen, it is necessary to predefine certain numbers of threshold values 1 (P1, P2, ...Pn) in order to provide different sensory perception zones. Each residence time within a zone and time interval between zones is recorded, as well as the elapse time between the transition from a lower to a higher threshold value and vice versa.
Fig. 2a shows the first of these three types of sensory time quantization. It is designated STQ(v) elapse time (i.e. sensitivity/time quantum of velocity), and produces a parameter for the relative moment speed vm. It can also be understood as the time duration between the phase transitions of two parallel signal traces at the same threshold value potential. That is similar to the standard term "phase shift". In the graph, the measured STQ(v) elapse times are designated with Tv(n). The phase transitions at the amplitude trace V, which is produced when the sensor (or receptor) 2 passes along a corresponding external signal source 4, are designated iTv(n.n); the phase transitions at the amplitude trace W, which are produced when the sensor (or receptor) 3 passes along the same signal source, are designated with iTw(n,n). In the ideal case, the sensors 3, 4 are close together compared to the distance c between external signal source and sensors, c remains approximately constant, and both sensors (or receptors) display identical properties and provide an analogue signal; then two amplitude traces V and W are produced at the outputs of the mentioned sensors (the sensor amplifiers or receptors, respectively) which are approximately congruent. (Deviations from ideal conditions are compensated by an autonomous adaptation of the sensory system in a continuously improved way, which is described later). When sensor 2, in the designated direction, moves along the signal source 4, then the signal amplitude V passes through the predefined threshold potential P1 at phase transition iTv(1.1). The rising signal edge actuates a first timer that records the first STQ(v) elapse time Tv(1). The continually rising signal amplitude V passes through the threshold potentials P2, P3 and P4; the phase transition of each of these activates further timers used for recording of further elapse times Tv(2), Tv(3) and Tv(4). Meanwhile, sensor 3 has approached signal source 4 and produces the signal amplitude trace W. When W passes through the threshold potential P1 at the phase transition iTw(1.1), the rising signal edge stops the timer, and the first STQ(v) elapse time is recorded and stored. The same procedure is repeated for the elapse times Tv(2), Tv(3) and Tv(4), when the signal amplitude passes through the next higher threshold values P2, P3 and P4. If V begins to fall, it first passes through the threshold value P4 on the falling shoulder of the amplitude trace. Now, the falling signal edge activates a timer that records the next elapse time Tv(5). At the further phase transitions iTv(3.2) and iTv(2.2), where the threshold values P3 and P2 are passed downwards, there are also timers which are actuated when the signal edges fall, in order to measure the elapse times Tv(6), Tv(7). If the signal amplitude V rises again, the STQ(v) parameters are recorded by the rising signal edges again. The same procedure is applied to stopping the timers at the phase transitions of the signal amplitude W. This produces the time displacement.
Fig. 2b shows another type of sensory STQ
quantization. It is called STQ(i) elapse time (i.e. sensitivity/time
quantum of interarrival). Simply, it is the time Tw a mobile system
needs for a relative length. It can also be understood as the time
duration between phase transitions of a signal trace at same
threshold potentials. If the time counting frequencies corresponding
to the relative speed parameters Tv, (i.e., the STQ(v) elapse times)
are proportionally accelerated or decelerated, the recorded
modulated time pulses correlate with the relative lengths. With
absolute physical invariance between the sensor and the signal
sources (i.e., synchronism), no STQ(v) parameter can be acquired,
but if an equivalent signal intensity is changing, STQ(v) data are
even obtainable when there is no relative motion. Therefore, during
motion, these data are necessary not only for recording variable
signals, but also for scanning spatial surroundings. In this figure,
measured STQ(i) elapse times are designated with Tw(n). The phase
transitions, which are produced by the amplitude trace W when the
sensor (or receptor) 5 is moving along the
corresponding adjacent signal sources 6 and
7, are designated with iTw(n.n). As soon as the sensor (or
receptor) 5 passes in the marked direction along the signal source
6, the signal amplitude W goes through the
pre-defined threshold potential P1 at phase transition iTw(1.1). The
rising signal edge activates a first timer for the recording of the
first STQ(i) elapse time Tw(1). Thereafter, the continually rising
signal amplitude W passes through the pre-defined threshold
potentials P2, P3 and P4, and when these show a phase transition,
further timers are activated in order to record further elapse times
Tw(2), Tw(3) and Tw(4). Meanwhile, sensor 5 begins
to move away from the vicinity of the signal source 6. The falling
amplitude trace passes through the threshold potential P4, and upon
the phase transition iTw(4.2) the falling signal edge stops the
timer that was recording the STQ(i) elapse time Tw(4).
Simultaneously, the same falling signal edge activates another timer
which measures the elapsed time Tw(5) up to the arrival of the next
rising signal edge. But this signal edge rises when sensor 5
passes along the equivalent signal source 7.
However, previously, the signal amplitude falls under the threshold
values P3 and P2, and when these show the phase transitions iTw(3.2)
and iTw(2.2), the timers recording the STQ(i) elapse times Tw(3) and
Tw(2) are stopped. Simultaneously, additional timers recording the
elapse times Tw(6) and Tw(7) are activated. They stop again at the
phase transitions iT(2.3), iTw(3.3), iTw(4.3) and iTw(5.1), when the
signal amplitude goes upwards again (but not before the sensor
motion along signal source 7 starts). After those phase transitions,
new timers start recording the next elapse times Tw(8), Tw(9),
Tw(10), Tw(11), and so on
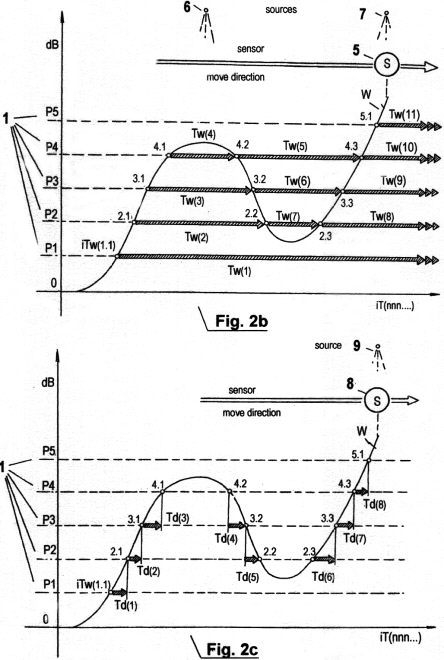
Fig. 2c shows a third type of sensory STQ
quantization that is completely different to those of Figs. 2a and
2b. It is termed STQ(d) elapse time (i.e., sensitivity/time quantity
of differentiation); and it can be understood as the time duration
Td, measured between a first phase transition at a first predefined
threshold potential up to the next phase transition at the next
threshold potential, which can be either higher or lower than the
first one. STQ(d) elapse times are parameters for the slope of
signal amplitude traces, and consequently they are parameters for
their frequency. By fast comparison of STQ(d) elapse times, signal
courses can be recognized in real time; therefore, for the creation
of intelligent behavior, STQ(d) quanta are just as imperative as
STQ(v) quanta and STQ(i) quanta. The quantization of STQ(d)-elapse
times is possible under all variable physical states and arbitrary
relative motion between sensor and external sources, in which STQ(v)
and STQ(i) elapse times are also quantizable. If the STQ(d) elapse
times are acquired cumulatively and serially, then they can be used
in the verification and plausibility examination of STQ(i) elapse
times (which are likewise acquired). In the graph, the measured
STQ(d) elapse times are designated with Td(n). The phase transitions
which are produced by the amplitude trace W when the sensor (or
receptor) 8 is in the field of a corresponding signal source 9, are
designated with iTw(n.n). When sensor 8 moves along the
corresponding signal-source 9 in the direction shown, at first the
signal amplitude W passes through the pre-defined threshold value P1
at the phase-transition iTw(1.1). Of course, this also happens when
the field of this signal source is active and/or variable, although
the sensor and the corresponding signal source are in an invariant
opposite position. The rising signal edge activates a first timer
that records the first STQ(d) elapse time Td(1). When the rising
amplitude trace W passes through the next higher threshold value P2
at the phase transition iTw(2.1), this timer is stopped and the
measured STQ(d) elapse time Td(1) is stored. Simultaneously, the
next timer is activated, and records the elapse time up to the next
phase transition at iTw(3.1), upon which it is stopped; then the
next timer is activated up to the next transition iTw(4.1), upon
which it is stopped again, and so on. (All the measured elapse times
are stored in memory). At the phase transition iTw(4.1) the next
timer is activated by threshold potential P4. However, since the
amplitude trace does not reach the next higher threshold value
before falling to P4 again, no STQ(d) can be acquired with the last
timer. Thus in this position only the quantization of STQ(i) elapse
times, as described in Fig. 2b, can take place. The next STQ(d)
elapse time Td(4) can only be acquired when the signal amplitude
falls below the threshold value P4 at the transition iTw(4.2), upon
which the next timer is activated, and stopped when the phase
transition at the next lower threshold value P3 occurs.
Simultaneously, the next timer is activated, and so on.
In
mechanistic applications, where the analysis of signal amplitudes
requires the quantization of STQ(d) elapse times, STQ(d) data are
often acquired in combination with STQ(i) data. If it is intended to
use this quantization method to enable a robot to recognize its own
motion from a subjective view (by detecting and scanning the spatial
surroundings when one moves along external signal sources), then
STQ(v) and STQ(i) data are predominantly acquired. However, if the
main intention is to recognize external, non-static optical or
acoustic sources such as objects, pictures, music or conversations
etc., then the proportion of STQ(d) parameters increases, while the
proportion of STQ(v) parameters decreases. In the case of physical
invariance (i.e. when there is no relative motion) no speed
parameters can be derived from any sensor signals, and only STQ(d)
and STQ(i) elapse times are quantized.
Figs. 3 a - c
illustrate an important aspect of the performance of the present
method, in connection with serial transfer of acquired STQ(d) elapse
times, as well as in connection with time pulse frequency modulation
in relation to simultaneously acquired STQ(v) parameters which
represent the instantaneous relative speed (vm). However, this
instantiation of the method is only suitable where mainly STQ(d)
elapse times are measured, together with those STQ(i) elapse times
(see also Fig. 2c) which are produced at the phase transitions when
maximal threshold value near the maximum of the amplitude are
reached, or when the minimal threshold value near the minimum of the
amplitude is reached. In this case, all measured elapse times can be
represented as serial data sequences. But if each phase transition
at each threshold potential generates STQ(d) elapse times as well as
STQ(i) elapse times (see also the notes for Fig. 5), then these data
are produced in parallel, and therefore they have to be processed in
parallel.
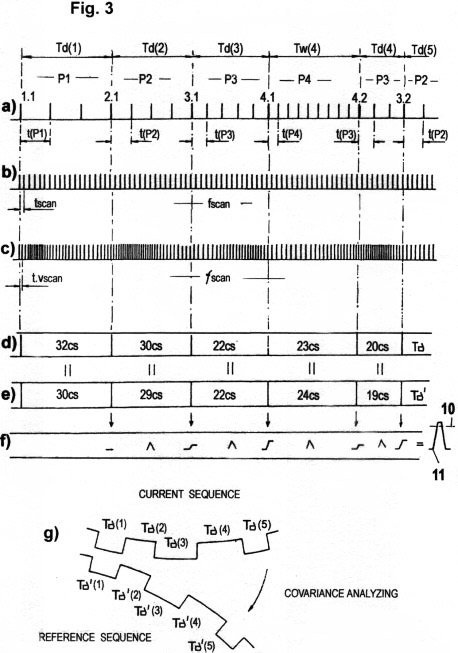
Fig. 3a shows how a simple serial pulse sequence can be sufficient for data transport of acquired STQ(d) elapse times, if the threshold potentials P1, P2, P3... that define the phase transitions 1.1, 2.1, 3.1... from which the STQ elapse times are derived, are "marked" either by codes or by certain characteristic frequencies. In this figure, these "markers" are pulses with period t(P1), t(P2), t(P3)... and frequencies f(P1), f(P2), f(P3).... These are modulated according to the respective threshold potentials. These identification pulses (IP) serve to identify the pre-defined threshold values P1, P2, P3...., (or the perception zones 1, 2, 3..., respectively). Only these identification pulses, in cooperation with invariable time counting pulses (ITCP) with the period tscan, or in cooperation with variable (vm modulated) time counting pulses (VTCP) with the period t.vscan (see also Figs. 3b, 3c), enable the actual acquisition of the STQ(d) elapse times Td(1), Td(2), Td(3), Td(4),... (or, respectively, the STQ(i) elapse times Tw(1), Tw(2), Tw(3), Tw(4),.... that are produced at amplitude maxima or minima), as we have already described. Variable VTCP pulses with the period t.vscan, which are automatically modulated relative to the acquired STQ(v) parameters (i.e., the instantaneous moment speed vm), are used to scan the signal amplitudes that are derived from external sources, in a manner proportional to speed. This reduces the redundancy of the calculation processes considerably (see also Fig. 3c). The STQ(d) elapse times that are acquired in such a vm-adapted manner by VTCP pulses are designated with Tδ; the STQ(i) elapse times, acquired in the same manner, are designated with Tω 1,2,3...).
Fig. 3b shows the
measurement of STQ(d) elapse times with invariant ITCP pulses with
period tscan and constant frequency fscan. This takes place as long
as no STQ(v) parameter is acquired, e.g. when no relative motion is
present between sensor and signal sources, and therefore when can be
measured.
Fig. 3c shows the
measurement of STQ elapse times with modulated VTCP pulses. These
counting pulses depend on the instantaneous relative speed vm (or on
the acquired STQ(v) parameter, respectively) as well as their period
t.vscan and frequency ƒscan in a manner that is proportion to vm. If
vm is very small or tends to zero, then the counting frequency ƒscan
is likewise reduced to the minimum frequency fscan (as seen in Fig.
3b). As shown in Fig. 2a, each STQ(v) parameter is acquired by means
of a second adequate "front" sensor (or receptor). Vm is thus
already recorded even before the actual STQ(d) and/or STQ(i) elapse
time measurement. Therefore it is possible automatically to modulate
ƒscan for the measurement of Tδ data according to the acquired
STQ(v) parameters, in order to reduce the number of t.vcalculations
as well as to minimize memory requirements. Thus, a largely
redundancy-free analysis results.
Although the time impulses
counted with this method are approximately covariant with the
covered lengths (d), it can be proved that they nevertheless
represent modified time data, and not distance data. As with the
origin of those data, the further processing and analysis of such
modified STQ elapse times Tδ (n) is dependent on probabilistic
principles. The time data T δ (n) are effectively "subjectively
sensed".
In mechanistic systems the modulation of time counting
frequencies in a manner proportional to distance traveled is done
chiefly by means of programmable oscillators and timers, as
illustrated in Fig. 5. However, in complex structured
biological/chemical organisms, this self-adaptive process (a part of
the so-called "autonomous adaptation") is generated mainly by
proportional >alteration of the propagation speed of timing pulses
in neural fibers, as shown in Figs. 4a -d. However, autonomous
adaptation and self-adaptive time base-altering processes of the
type described can also be formed differently. They can exist on
molecular, atomic or subatomic length scales. The author names this
principle "temporal auto-adaptation".
Figs. 3d - g show the
conceptual basis for the comparison of currently acquired STQ time
data sequences with prior recorded STQ time data sequences, as
well as their statistics-based analysis. The vm-modulated time
data Tδ (n), shown in Fig. 3d having the sequence 32 30 22 23 20 (cs
= cycles), are compared datum by datum with prior recorded time
data Tδ' (n), having the sequence 30 29 22 24 19, which were
likewise recorded in a vm-modulated manner. The comparison process
is actually a covariance analysis. When the regression curves of
both time data patterns converge, covariance exists. For these
purposes, in mechanistic systems, coincidence measurement devices,
comparator circuits, software for statistical analysis methods
or "fuzzy logic" can be used. The probability density
parameters are added up, and as soon as the total value within a
certain period exceeds a pre-defined threshold 10, then a signal
11 is produced that indicates that the sequence was "recognized".
This signal predominantly serves to regulate adaptively the
actuators in mechanistic systems (or motor behavior in organisms,
respectively). Moreover, the signal shows that "autonomous
adaptation" has taken place prior to these time data patterns being
recorded. In respect of the motoric behavior of any mechanistic or
biological organism, it is true that recognition of signal
sequences goes hand in hand with automatic adaptation (or
"autonomous adaptation", respectively). This principle is hereby
termed "motoric auto-adaptation" or "auto-emulation".
Fig. 3g shows this auto-adaptation process in a
schematic and easily comprehensible manner. A currently acquired
T time data sequence is continually compared with prior recorded
Ttime data sequences, and if approximate covariance appears, then
the sequences fit like a key into a lock. As described in the
following sections, this process produces a type of "bootstrapping"
or "motoric emulation, which constitutes a basic characteristic
of redundancy-free autonomous self-organizing systems and
organisms. Admittedly, the covariance analysis of two time data
patterns in mechanistic/ electronic systems is relatively
complicated (see also Fig.5). But this is not so in
molecular/biological organisms and other systems. In such systems,
this "bootstrapping" appears as a so-called "synergetic effect",
which is approximately comparable with rolling a number of
billiard balls into holes arranged in some pattern. (The name
"synergetic" was first used by H. HAKEN in the year 1970.)
Successful potting is determined by speed and direction. If the
speed and direction are altered, no potting will takeplace. An
attempt can also fail if the positions of the holes was somehow
changed whilst the initial positions of the balls were kept
constant, even if their speed and direction were covariant with the
original speed and direction (and when the covariance does not
adequately take into account the changing pattern). In a similar
way, a current STQ time data sequence, acquired by an autonomous
self-organizing system, produces a characteristic fingerprint
pattern, and whenever a previously recorded reference pattern is
detected that is isomorphic to the currently recorded pattern, then
auto-adaptation and auto-emulation results. This phenomenon is
inherent in all life forms, organisms and elementary structures
as a teleological principle. If no covariant reference pattern is
found, the auto-adaptive regulating collapses and the system
behaves chaotically. This motion changes from chaotic back to
ordered as soon as currently recorded STQ time patterns begin to
converge to prior recorded STQ time patterns that the analyzer
finds to be covariant.
Figs. 4a - d
illustrate a model for the acquisition and processing of STQ(d) and
STQ(v) elapse times (see also Figs. 3a-g) and for temporal and
motoric auto-adaptation in a molecular/biological context. The
basic elements of the model have already been described in the
neurophysiology literature by KATZ,GRAY, KELLY, REDMAN, J. ECCLES
and others. The present invention is of special originality
because temporal and motoric auto-adaptation is effected here by
means of STQ quanta, which are described for the first time here.
Such systems consist mainly of numerous neurons (nerve cells).
The neurons are interconnected with receptors (sensory neurons)
which enables the recording and recognition of the neurons' physical
surroundings. In addition, the neurons cooperate with effectors
(e.g.muscles) which serve as command executors for the motoric
activity. The expression "receptor" or "sensory neuron" corresponds
to the mechanistic term "sensor". An "effector" is the same as an
"actuator", which is a known term in the cybernetics literature.
Each neuron consists of a cell membrane that encloses the cell
contents and the cell nucleus. Varying numbers of branches from
the neurons (axons, dendrites etc.) process information off to
effectors or other neurons. The junction of a dendritic or axional
ending with another cell is called a synapse. The neurons
themselves can be understood as complex biomolecular sensors and
time pulse generators; the synapses are time data analyzers which
continually compare the currently recorded elapse time sequences
with prior recorded elapse time patterns that were produced by the
sensory neurons and were propagated along nerve fibers towards the
synapses. In turn, a type of "covariance analysis" is carried out
there, and adequate probability density signals are generated that
propagate to other neighboring neural systems or to effectors.
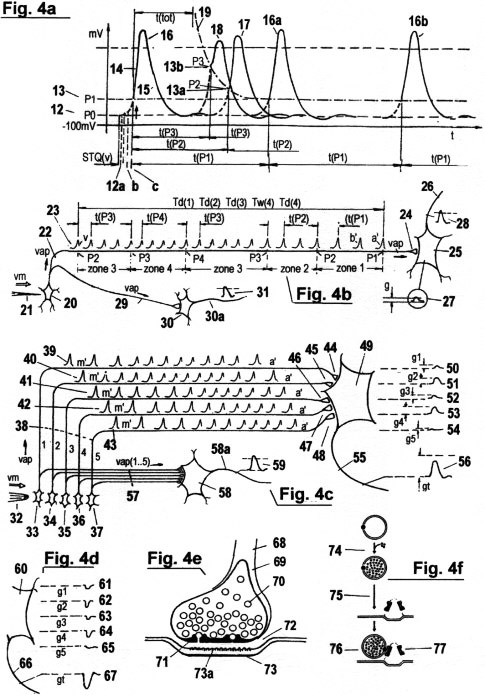
Fig. 4a shows a so-called "action potential" AP that is produced at the cell membrane by an abrupt alteration of the distribution of sodium and potassium ions in the intra and extra-cellular solution, which works like a capacitor. These ionic concentrations keep a certain balance as long as no stimulus is produced by the receptor cell. In this equilibrium state, a constant negative potential 12, termed the "rest potential", exists at the cell membrane. As soon as a receptor perceives a stimulus from an externalsignal source, Na+ ions flow into the neutral cell, which causes the distribution of positive and negative ions to be suddenly inverted, and the cell membrane " depolarizes". Depending on the intensity of the receptor stimulus, several effects are produced:
(a) If the threshold P1 is not exceeded, then a
so-called "electrotonic potential" EP is produced which propagates
passively along the cell membrane (or axon fiber), and which
decreases exponentially with respect to time and distance traveled.
The production of EP is akin to igniting an empty fuse cord. The
flame will stretch itself along the fuse, becoming weaker as it goes
along, before finally going out. EP's originate with each
stimulation of a neuron.
(b) If the threshold P1 is
exceeded, then an "action potential" AP (as in Fig. 4a) is produced
which propagates actively along the cell membrane (or axon fiber)
with a constant amplitude in a self-regenerating manner. The
production of AP is akin to a spark incident at a blasting fuse: the
fiercely burning powder heats neighboring parts of the fuse, causing
the powder there to burn, and so on, thus propagating the flame
along the fuse.
AP's are used in the quantization of STQ(d) and STQ(v) elapse times. They are practically equivalent to identification pulses IP with periods t(P1), t(P2), t(Pn)..., which are shown in Fig. 3a. AP's signal the occurrence of the phase transitions from which STQ(d) and STQ(v) elapse times derive. In addition,the AP' indirectly activate the molecular/biological "timers" that are used for recording these elapse times. But AP's do not represent deterministic sampling rates for amplitude scanning; and they do not correspond to electronic voltage/frequency converters. Moreover, their amplitude is independent of the stimulation intensity at the receptor, and they do not represent the time counting pulses used in the measurement of elapse times. Rather, the recording of STQ elapse times is effected and modulated by the velocity with which the action potentials propagate along the nerve fibers (axons) and membrane regions.
The time measuring properties of AP's are
described in detail in the following section:
If an EP, in answer
to a receptor stimulus, exceeds a certain threshold value (P1)
13, then an AP is triggered. The amplitude trace of
an AP begins with the upstroke 14 and ends with the
repolarisation 15, or with the so-called
"refractory period", respectively. At the end of this process, the
membrane potential decreases again to the resting potential P0, and
the ionic distribution returns to equilibrium. Not each receptor
stimulus generates sufficient electric conductivity to produce an
AP. As long as it remains under a minimal threshold value P1, it
generates only the electrotonic potential EP (introduced above).
(For a better understanding of elapse time measurements in
biological/chemical structures, see Fig. 2c and Fig. 3a). The first
AP, which is triggered after a receptor is stimulated, generates
initially (indirectly) the impulse that activates the first timer
that records the first STQ(d) elapse time, when the signal amplitude
W passes through the threshold value of the potential P1 at phase
transition iTw(1.1). This signal represents simultaneously an
identification pulse IP. The first AP corresponds to the first IP in
a sequence of IP's that represents the respective threshold value
status or perception zone in which the stimulation amplitudes were
just found. As long as the stimulus at the receptor persists, an AP
16a, 16b... is triggered in temporal intervals
whose duration depends on the respective thresholds in which the
stimulus intensities have just been found.
These temporal
intervals correspond to those IP periods t(P1), t(P2),... that are
required for serial allocation and processing of STQ elapse times
(see Fig. 3a). The AP frequency is stabilised through the so-called
"relative refractory period" (i.e. downtime) after each AP, during
which no new depolarisation is possible. Because the relative
refractory period shortens itself adaptively in proportion to the
increase in stimulation intensity at the receptor (e.g. if the EP
reaches a higher threshold value P2 (or perception zone) 13a), there
is a similarity here with "programmable bi-stable multivibrators"
found in the usual mechanistic electronics. The downtime (refractory
period) after an AP is shown as the divided line 19.
Fig. 4a illustrates an "absolute refractory period" t(tot) following a repolarisation. No new AP can be created during this time, irrespective of the stimulation intensity at the receptor rises. The maximum magnitude of a recognizable receptor stimulus is programmed in this way. Of importance is the fact that both the duration of the relative refractory period as well as character of the absolute refractory period are subordinate to auto-adaptive regularities, and are therefore continually adapting to newly appearing conditions in the organism. Consequently, the threshold values P0, P1, P2.... from which STQ quanta are derived are themselves not absolute values, but are subject to adaptive alteration like all other parameters; including, in particular, the physical "time".
We shall now elaborate upon what happens after the first STQ(d)
elapse time at P1 is recorded via the first AP: If the stimulation
intensity (with a theoretical amplitude W) increases from the lower
threshold P1 to the next higher threshold P2, then the following AP
triggers indirectly the recording of the second STQ(d) elapse time
as soon as a phase transition occurs through the next higher
threshold P2. The same process is repeated in turn for the threshold
values P3, P4, ... and so on. In each case, the AP functions
simultaneously as an identification pulse IP, as described in Fig.
3a. It therefore recurs in threshold-dependent periods as long as a
perception acts upon the receptor (i.e. for as long as the receptor
is perceiving something).
As an example, consider also Fig.
3a: As long as the stimulation intensity remains in the zone P2, the
AP 17, 17a, 17b.... recurs in short temporal
periods. These periods (or intervals) are similar to those periods
of IP identification pulses (with period t(P2)) that are required
for serial recording of the STQ elapse times Td(2) and Tw(2). When
the increasing stimulation intensity reaches the threshold value P3
(or perception zone 3) 13b, the AP's recur in even shorter time
periods 18a, 18b, 18c... This corresponds to the IP identification
pulses with the period t(P3), shown in the figure, which are
indirectly required for serial timing of the STQ elapse times Td(3)
and Tw(3). An even larger stimulation intensity, for example in P4
(perception zone 4), would generate an even shorter period for the
AP's. This would correspond approximately to t(P4) in Fig. 3a. The
maximum possible AP pulse frequency is determined by t(tot). Shorter
refractory periods, after the depolarization of APs, also produce
smaller AP-amplitudes. This property simplifies the allocation of
AP's in addition. In the following, the generation of the actual
time counting pulses for STQ quantization is detailed. These pulses
are either invariable ITCP or vm-proportional VTCP, as illustrated
in Fig. 3a. The time counting pulses for the quantization of elapse
times are dependent on the velocity with which the AP propagate
along an axon. This velocity is in turn dependent on the "rest
potential" and on the concentration of Na+ flowing into the
intracellular space at the start of the depolarization process, as
soon as perception at the receptor cell causes an electric current
to influence the extra/intra-cellular ionic equilibrium.
With the commencement of stimulation of a receptor (at the outset of a perception), only capacitive current flows from the extra-cellular space into the intracellular fluid. This generates an "electrotonic potential" EP, which propagates passively. If this EP exceeds the threshold P1, then an AP, which propagates in a self-regenerating manner along the membrane districts, is produced. The greater the capacitive current still available after depolarisation (or "charge reversal") of the membrane capacitor, the greater the Na+ ion flow into the intracellular space, and the greater the available EP current that can flow into still undepolarized areas. The rate of further depolarization processes in the neuronal fibres, and consequently the propagation speeds of further AP's, are thus increased proportionally. The charge reversal time of the membrane capacitor is therefore the parameter that determines the value 12 of the resting potential P0. When a stimulus ("excitation") starts from the lowest resting potential 12, then the Na+ influx is the largest, the EP-rise is steepest and the electrotonic flux is maximum. If an AP is triggered, then its propagation speed is in this case also maximum. But when a receptor stimulus starts from a higher potential 12a, 12b, 12c...., then the Na+ influx is partially inactivated, and the steepness of the EP-rise as well as its electrotonic flux velocity is decreased. Therefore, the propagation speed of an AP decreases too.These specific properties are used in molecular/biologic organisms to produce either invariant time counting impulses ITCP, with periods tscan, or variable time counting impulses VTCP with periods t.vscan. In the latter case, the VTCP's are modulated in accordance with the relative speeds vm (via the STQ(v) parameters), and therefore have shorter intervals (see Figs. 3b, 3c). The STQ(v)-quantum is determined by the deviation of the respective starting-potential from the lowest resting-potential P0, which serves as a reference value, and is measured by the duration of the capacitive charging of a cell membrane when a stimulus occurs at the receptor.
The duration of the charging is inversely proportional to the
velocity of the Na+ influx through the membrane channels into the
intracellular space. A cell membrane can be understood as an
electric capacitor, in which two conducting media, the intracellular
and the extracellular solution, are separated from one another by
the non-conducting layer, the membrane. The two media contain
different distributions of Na/K/Cl ions. The greater the
"stimulation dynamics" (see below) that first influences the outer
molecular media - corresponding to sensor 2 in Fig.
2a - and, subsequently, the inner molecular media - which
corresponds to sensor 1 in Fig. 2a - the faster is
the Na+ influx and the shorter the charging time (which determines
the parameter for the relative speed vm), and the faster is the AP
propagation velocity v(ap) in the neighbouring membrane districts.
The signals at the inner and outer sides, respectively, of the
membrane, correspond to the signal amplitudes V and W. The velocity
v(ap), therefore, indirectly generates the invariant time counting
pulses ITCP or the variable vm-proportional time counting pulses
VTCP.
These variable VTCP pulses are self-adaptive modulated time pulses that are correlated to the relative length. As explained in the following (contrary to the traditional physical sense), no "invariant time" exists -- only "perceived time" exists. Of essential importance also is the difference between "stimulation intensity " whose measurement is determined by the AP frequency and therefore by the refractory period, and the "stimulation dynamics", whose measurement is defined by the charge duration of the cell membrane and therefore also by the speed of the Na+ influx. "Stimulation dynamics" is not the same as "increase of the stimulation intensity". It is a measure of the temporal/spatial variation of the position of the receptor relative to the position of the stimulus source, and therefore of the relative speed vm. The stimulation intensity corresponds to signal amplitudes, from which vm-adaptive STQ(d) elapse times Tδ(1,2,3...) are derived, while the stimulation dynamics is defined by the acquired STQ(v) parameters. `
Fig. 4b and Fig. 4c show the
analysis of STQ elapse times in a molecular/biological model in an
easily comprehensible manner. The results of the analysis are used
to generate redundancy-free auto-adaptive pattern recognition as
well as autonomous regulating and self-organization processes. The
organism in the particular example shown here is forced to
distinguish certain types of foreign bodies that press on its
"skin". It must reply with a fast muscle reflex when it recognizes a
pinprick. But it should ignore the stimulus when it recognizes a
blunt object. A continuous vm-adaptive recording of STQ(d) elapse
times by means of VTCP pulses is necessary to do this. The frequency
of these time counting impulses is modulated in accordance with the
STQ(v) parameters of the stimulus dynamics (vm). These STQ(v)
parameters are required for the recording of the STQ(d) elapse times
Tδ(1,2,3...) from the signal amplitude at the current stimulus
intensity. The difference between "stimulation intensity" and
"stimulation dynamics" is easily seen in this example. A stimulus
can even show a different intensity if no temporal-spatial change
takes place between signal source and receptor. A needle in the skin
can cause a different sensory pattern even when its position is not
changing if, for example, it is heated. This sensory pattern is
determined by the signal amplitude, and consequently by the AP
frequency and by the STQ(d) quanta. As long as the needle persists
in an invariant position, the AP propagation velocity is constant,
because the membrane charging time is constant too. During the prick
into the skin, there is a "dynamic stimulation", and the STQ(d)
quantization of the signal amplitude is carried out in a manner that
depends on the pricking speed vm. It should be noted that two
temporally displaced signal amplitudes (at the inner and outer
membrane surface) always exist during this dynamic process. The
STQ(v) parameters are derived from this. The AP propagation
velocities and the acquired STQ(d) time patterns are adapted
accordingly ("temporal auto-adaptation").
The STQ(d) time
patterns Tδ(1,2,3,4,.....), measured adaptively according to the vm,
are constantly compared to and analysed together with the previously
measured and stored STQ(d) time patterns Tδ'(1,2,3...). This time
comparation process occurs continuously in the so-called synapses,
which are the junctions to axional endings of other neurons. The
probability density values that are produced at the synapses, and
which are used to represent the convergence of both regression
curves, are communicated for further processing to peripheral neural
systems, or to muscle fibres in order to trigger motoric reflex.
Fig. 4b shows the vm-dependent propagation of an AP from a sensory neuron (receptor) 20 along an axon to a synapsis, where a comparison of acquired time sequences takes place through molecular" covariance analysis". This receptor functions like a "pressure sensor". If a needle 21 with a certain dynamics impinges on the outer side of the cell membrane, then this stimulation causes triggering of AP's 23 as described in Fig. 4a. The AP's propagate in the axon 22 with a STQ(v)-dependent speed vap. The sequence (a'.....v') represents the signal amplitude values that are produced by the pinprick. The sequence begins with the phase transition at the first threshold value P1, continues over P2, P3, P4 (at which point the stimulus maximum is attained), and finally to the phase transitions through P3 and P2. The intensity zones for stimulus perception are designated with Z1, Z2, Z3 and Z4. The periods t(P1), t(P2), t(P3), t(P4)......, and the magnitudes of the AP's serve to identify the particular threshold in which the stimulation intensity is currently to be found. Their temporal sequence is therefore a type of "code". AP's are not time counting pulses. Besides their coding function, they also serve as (indirect) activating and deactivating pulses for the recording of STQ(d) elapse times. The actual vm-dependent measurement of the STQ elapse times Td(1), Td(2), Td(3), Tw(4) and Td(4)... (see Fig. 2c), as well as the comparison of these with previously recorded elapse times, takes place in the synapse 24. At the presynaptic terminal of the axons, the AP's 23 arrive with variable velocities vm(n...), according to the dynamics of the needle prick as well as the measured STQ(v) parameters. This variable arrival velocity at the synapses is the key to producing the adaptive time counting impulses VTCP (see Fig. 3c) with vm-modulated frequency ƒscan. The synapse is separated from the postsynaptic membrane by the "synaptic cleft", and the postsynaptic membrane, for its part, is interconnected with other neurons; for instance, to a "motorneuron" 25. This neuron generates a so-called "excitatory postsynaptic potential" (ESPS) 27 that is approximately proportional to the convergence probability g. If this EPSP (or, equivalently, the probability density g) exceeds a certain threshold value, then, in turn, an action potential AP 28 is triggered. This AP is communicated via motoaxon 26 to the "neuromuscular junction", at which a muscle reflex is triggered. The incoming AP sequences 23 generate the release of particular amounts of molecular transmitter substance from their repositories - tiny spherical structures in the synapse, termed "vesicles". In principle, a synapse is a complex programmable timedata processor and analyzer that empties the contents of a vesicle into the presynaptic cleft when the recurrence of any prior recorded synaptic structure is confirmed within a newly recorded key sequence. The synaptic structures and vesicle motions are generated by the dynamics (vap) of the AP ionic flux, as well as by its frequency. AP influx velocities v(ap) correspond to the STQ(v) elapse times, and AP frequencies correspond to the STQ(d) elapse times. The transmitter substance is reabsorbed by the synapse, and reused later, whereby the cycle continues uninterrupted.
We now present a detailed description of Fig. 4b (referring
also to Figs. 4e and 4f). The ionic influx of the initial incoming
AP 23 (a') activates the spherical structures
(vesicles) containing the ACh transmitter molecules. These molecules
are released in the form of a "packet". The duration of this ACh
packaging depends on the dynamics (represented by the velocity
v(ap)) of the AP ionic influx at the presynaptic terminal, and
therefore on the stimulus dynamics (represented by vm) at the
receptor 20. Each subsequent incoming AP, namely
b', c'..., in turn causes neurotransmitter substances in the vesicle
to be released toward the synaptic cleft. Each of the following are
elapse time counting and covariance analyzing characteristics:: the
duration of accumulation of neurotransmitter substance T(t); the
velocities v(t) with which the neurotransmitter substances move in
the direction of the synaptic cleft; the effects induced by the
neurotransmitter substances at the synaptic lattice at the synaptic
cleft; the duration of pore opening; and so on. By means of AP's
acting on synaptic structures, not only are the actual time counting
frequencies ƒscan generated (to be used in vm-dependent measurement
of STQ(d) elapse times as described in Fig. 2c), but also time
patterns are stored and analysed.
If the pattern of a current temporal sequence is recognised by the synapse as matching an existing stored pattern, a pore opens at the synaptic lattice, and all of the neurotransmitter content of a vesicle is released into the subsynaptic cleft. The released transmitter molecules (mostly ACh) combine at the other side of the cleft with specific receptor molecules of the sub-synaptic membrane of the coupled neuron. Thus, a postsynaptic potential (EPSP) is generated, which then propagates to other synapses, dendrites, or to a "neuromuscular junction". If the EPSP exceeds a certain amplitude, then it triggers an action potential (AP) of the described type, which then triggers, for example, a muscle reflex. If the potential does not reach this threshold, then the EPSP propagates in the same manner as an EP (i.e. in an electrotonic manner); an AP is not produced in this case.
Of special significance is the summing property of the
subsynaptic membrane. This characteristic, termed "temporal
facility", results in the summation of amplitudes of the generated
EPSP's, if they arrive in short sequences within certain time
intervals. Each release of neurotransmitter molecules into the
synaptic cleft designates an increased probability density occurring
during the comparison of instantaneous vm-proportionally acquired
STQ time patterns to prior vm-proportionally recorded STQ-time
patterns. Increased probability density causes a higher frequency of
transmitter substance release and therefore a higher summation rate
of the EPSP's, which in turn produces, at a significantly increased
rate, postsynaptic action potentials (AP). Therefore, a postsynaptic
AP is effectively a confirmation signal that flags the fact that
isomorphism between a previously and currently recorded time data
pattern has been recognized. On the basis of this time pattern
comparison, the object that caused the perception at the receptor
cell is thereby identified as "needle"; and the command to "trigger
a muscle reflex" is conveyed to the corresponding muscle fibres.
Parallel and more exact recognition processes are executed by the
central nervous system CNS (i.e. the brain). From the sensitive
skin-receptor neuron 20, a further axonal branching
29 is connected via a synapse 30
to a "CNS neuron". In contrast to the "motorneuron" which actuates
the motoric activity of the organism directly, a CNS neuron serves
for the conscious recognition of a receptoric stimulation sequence.
An AP 31, produced at the postsynaptic cell
membrane 30, can spread out along dendrites in the
axon 30a, as well as to several other CNS neurons;
or, alternatively, indirectly via CNS neurons to a motorneuron, then
on to a neuromuscular junction.
The parameters controlling
the recording of STQ time quanta in the synapses 25
and 30 can differ with different synaptic
structures. (Indeed, the synaptic structures themselves are
generated by continuous "learning" processes). This explains how it
is possible for a needle prick to be registered by the brain, while
eliciting no muscular response; or how a fast muscle reflex can be
produced while a cause is hardly perceived by the brain. The first
case shows a conscious reflex, the other case an instinctive reflex.
The former occurs when the CNS synapse 30 cannot
find enough isomorphic structures (in contrast to the synapse
25), transmitter molecules are not released with
sufficient frequency, and subsequently no postsynaptic AP 31 and no
conscious recognition of the perceived stimulus can take place.
Numerous functions of the central nervous system can be explained in
such a monistic way; as well as phenomena such as "consciousness"
and "subconscious". Generally, auto-adaptive processes are deeply
interlaced in organisms, and are therefore extremely complex. In
order to be capable of distinguishing a needle prick from the
pressure of a blunt eraser, essentially more time patterns are
necessary; in addition, more receptors and synapses must be involved
in the recognition process.
Fig. 4c
illustrates the process by which moderate pressure from a blunt
object (e.g. a conical eraser on a pin) is recognized, resulting in
no muscle reflex. The blunt object 32 presses down
with a certain relative velocity vm onto a series of receptors in
neural skin cells 33, 34, 35, 36 and 37.
Several sequences of AP's 39, 40, 41, 42 and
43 are produced after the individual adjacent
receptors (see also Fig. 4b) are stimulated. These action potentials
propagate along the collateral axons 38 with
variable periods t(P1,2,3..) and velocities vap(1..5), which result
on the one hand from the prevailing stimulation intensity, and on
the other hand from the respective stimulation dynamics. Since each
receptor stimulus generates a different pattern of STQ(v) and STQ(d)
quanta, various AP sequences a'.....m' emerge from each axon. All
sequences taken together represent the pattern of STQ elapse times
which characterises the pressure of the eraser on the skin. These
variable AP ionic fluxes reach the synapses 44, 45, 46, 47
and 48, which are interconnected via the synaptic
cleft with the motoneuron 49. As soon as the
currently acquired STQ time data pattern shows a similarity to a
prior recorded STQ time data pattern, each individual synapse
releases the contents of a vesicle into the subsynaptic cleft.
Simultaneously, this produces an EPSP at the subsynaptic membrane of
the neuron. These EPSP potentials are mostly below the threshold.
The required threshold value for the release of an AP is reached
only when a number of EPSP's are summed. This happens only when a
so-called "temporal facilitation" of such potentials occurs, as
described in the previous paragraph.
In the model shown, the
individual EPSP's 50, 51, 52, 53 and 54
effect this summing property of the subsynaptic membrane. These
potentials correspond to receptor-specific probability density
parameters g1, g2, g3, g4 and g5, that represent the degree of
isomorphity of time patterns. Simultaneous neurotransmitter release
in several synapses, for example in 45 and
47, causes particular EPSP's to be summed to a total
potential 56, which represents the sum of the
particular probability densities G = g1+g3. This property of the
neurons (i.e. the summing of spatially separated subliminal EPSP's
when release of neurotransmitter substance appears simultaneously at
a number of parallel synapses on the same subsynaptic membrane) is
termed "spatial facilitation".
In the described model case,
the summed EPSP 56 does not, however, reach the
marked threshold (gt), and therefore no AP is produced. Instead, the
EPSP propagates in the sub-synaptic membrane region 49
of the neuron, or in the following motoaxon 55,
respectively, as a passive electrotonic potential (EP). Such an EP
attenuates (in contrast to a self-generating active AP) a few
millimetres along the axon, and therefore has no activating
influence on the neuromuscular junction, and consequently no
activating influence on the muscle. The stimulation of the skin by
pressing with the eraser is therefore not sufficient to evoke a
muscle reflex.
It would be a different occurance if the eraser
would break off and the empty pin meet the skin receptors with full
force. In this case, neurotransmitter substances would be released
simultaneously in all five synapses 50, 51, 52, 53
and 54, because the acquired STQ time patterns
Tδ(1,2,3..), with very high probability, would be similar to those
STQ time patterns Tδ'(1,2,3... ) already stored in the synaptic
structures that pertain to the event "needle prick". The EPSP's
would be summed, because of their temporal and spatial
"facilitation", to a supraliminal EPSP 56, and a
postsynaptic AP would be produced that propagates along the motoaxon
55 in a self-regenerating manner (without temporal
and spatial attenuation) up to the muscle, producing a muscle
reflex.
As in Fig. 4b, in the present example a recognition process takes place in the central nervous system (CNS) that proceeds in parallel. From the skin receptor cells 33, 34, 35, 36 and 37, collateral axonal branches extend to CNS synapses that are connected to other neurons 58. Such branches are termed "divergences". The subdivision of axons into collateral branches in different neural CNS districts, and the temporal and spatial combination of many postsynaptic EPSP's, allows conscious recognition of complex perceptions in the brain (for example, the fact of an eraser pressing onto the skin). Since this recognition has to take place independent of the production of a muscle reflex, the sum of individual EPSP's must be supraliminal in the CNS. Otherwise, no postsynaptic AP - i.e. no signal of confirmation - can be produced. As an essential prerequisite for this, it is necessary that auto-adaptive processes have already occurred which have formed certain pre-synaptic and sub-synaptic STQ time structures in the parallel synapses 58. These structures hold information (time sequences; i.e. patterns) pertaining to similar sensory experiences (e.g. "objects impinging on the skin" - amongst these, a conical eraser). Obviously the threshold for causing an AP in the postsynaptic membrane structure of the ZNS Neurons 58 (and therefore also in the brain) has to be lower than in the motoneuron membrane 49 described previously. Therefore also the sum of these EPSP's must be larger than the sum of the EPSP's g1, g2, g3, g4 and g5. Isomorphisms of STQ time patterns in the CNS synapses of the brain have to be more precisely marked out than those in the synapses of motoneurons, which are only responsible for muscle reflexes
The structure of the CNS synapses must be able to discern finer
information, so it must be more subtle. The production of a
sub-synaptic AP represents a confirmation of the fact that a
currently acquired Tδ(1,2,3...) time pattern is virtually isomorphic
to a prior recorded reference time pattern Tδ'(1,2,3...), which, for
example, arose from a former sensory experience with an eraser
impinging at a certain location on the skin. If such a former
experience has not taken place, the consciousness has no physical
basis for the recognition, since the basis for time pattern
comparison is missing. In such a case, therefore, a learning process
would first have to occur. Most of the time, however, sensory
experiences of a visual, acoustic or other type, arising from a
variety of receptor stimulation events, are co-ordinated with the
pressure sensing experience.
This explains why CNS structures
are extremely intensively interlaced. CNS neurons, as well as
moto-neurons, have up to 5000 coupled synapses, which are
interconnected in a multifarious manner with receptor neurons and
axonal branches. There are complex time data patterns for lower and
higher task sites, which are structured in a hierarchical manner. We
have already described simple Tδ(1,2,3....) and Tδ'(1,2,3...)
analysis operations. Blood circulation, respiration, co-ordination
of muscle systems, growth, seeing, hearing, speaking, smelling, and
so on, necessitate an extremely large number of synaptic recorded
"landscapes" of the organism's STQ time patterns, produced by a
variety of receptors; and which continually have to be analysed for
isomorphism with time patterns currently being recorded.
Accordingly, temporal and motoric auto-adaptation occurs in deeper
and higher hierarchies and at various levels.
Fig. 4d illustrate the counterpart to the EPSP
(Excitatory Postsynaptic Potential): the "Inhibitory
Postsynaptic
Potential " , or IPSP. As seen in the figure, the IPSP potentials
61, 62, 63, 64 and 65 at the
subsynaptic membrane 60 are negative compared to
the corresponding EPSP's. IPSP's are produced by a considerable
proportion of the synapses to effect pre-synaptic inhibition instead
of activation. The example here shows an IPSP packet 67
propagating from the motoaxon 66 to a neuromuscular
junction (or muscle fibre, respectively) which prevents this muscle
from being activated - even if a supraliminal EPSP were to reach the
same muscle fibre at the same time via a parallel motoaxon.
Positive EPSP's ion fluxes and negative IPSP's ion fluxes
counterbalance each other. The main function of the IPSP's is to
enable co-ordinated and homogeneous changes of state in the
organism, e.g. to enable exact timing of motion sequences. In order
to ensure, for example, a constant arm swing, it is necessary to
activate the bicep muscles, which then flex the elbow with the aid
of EPSP's; but to inhibit the antagonistic tricep muscles (which
extend the elbow) with the aid of IPSP's. Antagonist muscles must be
inhibited via so-called "antagonistic motoneurons", while the other
muscle is activated via "homonym motoneurons". The complex synergism
of excitatory (EPSP) synapses and inhibitory (IPSP) synapses act
like a feedback system (servoloop) and enables optimal timing and
efficiency in the organism. One can compare this process with a
servo-drive, or with power-steering, which ensures correct
co-ordination and execution of current motion through data-supported
operations and controls. If data are missing, the servoloop
collapses. Disturbances in a molecular biological servoloop that is
supported by STQ time data structures lead to tetanic twitches,
arbitrary contractions, chaotic cramps and so on.
From the
point of view of cybernetics, each excitatory synapse generates a
"motoric impulse" (EPSP), while each inhibitory synapse generates a
"brake impulse" (IPSP). The continued tuning of the complicated
servoloops, and the balance which results from continuous comparison
of prior sensory experiences (the stored reference time patterns)
with current sensory experiences (the time patterns currently being
recorded), creates "perfect timing" in the organism.
Fig. 4e shows the basic construction of a synapse. Axon
68 ends at the pre-synaptic terminal 69,
which is also termed "bouton". The serial incoming AP's cause the
vesicles to be filled with neurotransmitter molecules. When the
filling process is finished, the vesicles begin to move in the
direction of the pre-synaptic lattice 71. If a
currently acquired time pattern is approximately isomorphic to an
existing time pattern (see also Fig. 4b), then a small canal opens
at an attachment site on the lattice, which releases the entire
contents of the vesicle into the narrow synaptic cleft 72.
This process is termed "exocytosis". The sub-synaptic neural
membrane 73 supports specific molecular receptors
73a, to which the released transmitter molecules
bind themselves. For a certain period, a pore opens, through which
the transmitter substance diffuses. The conductivity of the
postsynaptic membrane increases and the EPSP (following postsynaptic
depolarisation) is triggered. The duration of opening of the pores
and the recognition of complementary receptors by the molecules are
likewise determined by auto-adaptive processes and evaluation of STQ
time pattern structures. However, these molecular processes
represent deeper sub-phenomena in comparison to synaptic processes.
Structures for temporal and motoric auto-adaptation, which depend on quantization of STQ-elapse times, also exist at the molecular and atomic levels.
Fig. 4f shows the filling of a vesicle
70 with neurotransmitting substances, and its subsequent
motion towards a pre-synaptic dense projection at the lattice
71. The start of the filling process 74
can be seen as the activation of a stopwatch. The rate v(t) of the
filling is proportional to the dynamics of the AP ionic flux into
the synapse. The periods T(t...) of the filling follow the periods
t(P1,P2,...) of the arriving AP's; these times, therefore, represent
vm-adaptive quantized STQ(d) elapse times Tδ(1,2,3...). The
direction of filling is shown at 75. The direction
of motion of a vesicle is shown at 76. If the
current velocity v(t), the duration of the vesicle packaging T(t),
the quantity of transmitter molecules, the current vesicle motion
and other currently significant STQ parameters have characteristics
which correlate to an existing synaptic STQ structure, then a filled
vesicle binds itself onto an "attachment site" 77
at the lattice. Ca++ ions flow into the synapse, a pore at the
para-crystalline vesicle lattice opens, and the entire molecular
neurotransmitter content is released into the synaptic cleft
72. At the postsynaptic membrane of the target neuron,
these molecules are fused with specific receptor molecules. Such
receptors have verification tasks. They prevent foreign transmitter
substances (that originate from other synapses) from producing wrong
ESPS's at this neuron.
To complete the discussion of Fig. 4,
we relate the descriptions of Figs. 4a, 4b, 4e and 4f to the
STQ-configurations of Figs. 3a - g. For argument's sake, we assume
once again that a pinprick impinges onto a receptor cell (see also
Fig. 4b). The IP sequences shown in Fig. 3a correspond to the AP's
23 which are produced by stimulating a receptor
cell 20 with a needle 21. Their
periods t(P1), t(P2),... serve to classify the respective zones of
stimulation intensity (P1, P2...) or perception intensity (Z1, Z2...
). Each AP 23, arriving into a synapse 69,
activates the adaptive quantization of STQ(d) elapse times,
depending on the velocity vap of the propagation of the AP along the
axon. Elapse timing with modulated time base is triggered as soon as
a vesicle begins to fill. Finished filling (packaging) signifies
"elapse timing stop, STQ(d)- quantum recorded". The elapse times
Td(1), Td(2), Td(3), Td(4).... thus recorded generate the
significant synaptic structures. Invariant time counting pulses ITCP
(see Fig. 3b) with frequency fscan correspond to constant axonal AP
propagation with velocity vap, if no dynamic stimulus appears at the
skin receptor cell (for example, if a needle remains in a fixed
position and generates a constant stimulation intensity). In this
case, the receptor membrane senses no relative speed vm; the AP's
propagate with constant velocity vap along the axon 22;
and the synapse quantizes the STQ(d) elapse times with invariant
time counting frequency fscan.
Time counting pulses VTCP (see
Fig. 3c) with variable frequency ƒscan are then applied, if dynamic
stimulation affects the receptor. The AP's propagate along the axon
with STQ(v)-dependent velocities vap(n...), modulated by the
variable dynamics vm(n...) which are measured as an STQ(v) parameter
by the membrane. Adaptive alteration of all of the following
processes occurs in a similar manner: the variation of time counting
periods t(P1... .n) corresponding to the points 2.1, 3.1, 4.1 in
Fig. 3c; the velocities v(t....) of AP ionic flux into the synapse;
the vesicle filling times T(t...); the amounts of transmitter
molecules contained in the vesicles; the motion of these molecules
in the direction of the vesicle lattice; the structure of this
lattice; and many other parameters of the presynaptic and
subsynaptic structures.
A synapse has features that enable
the conversion of the AP influx dynamics into vap-proportional
molecular changes of states. This is like the variable VTCP time
counting pulses seen in Fig. 3c. The process can be compared with
variable water pressure driving a turbine, through which a generator
produces variable frequencies depending on pressure and water speed:
higher water pressure is akin to higher stimulation dynamics vm at
the receptor, higher AP propagation velocity vap along the axon, and
higher VTCP time pulse frequency ƒscan in the synapse (which in turn
affects not only the rate v(t) with which vesicles are filled, but
also many other synaptic parameters). According to these processes,
the STQ(d) time sequence Td(1, 2, 3, 4...) is recorded in the
synapse with vm-modulated time counting frequencies ƒscan(1,2,3...);
as a consequence, the physical structure of the synapse is
determined by this time sequence.
Fig. 3d shows a currently
acquired time data sequence 32 30 22 23 20 that is
equivalent to the recorded time pattern Tδ(1,2,3..), and which
leaves a specific molecular biological track in the synapse
24. The prior acquired time data sequence 30 29 22
24 19 in Fig. 3e corresponds to the synaptic structure that
has been "engraved" through frequent repetition of particular
stimulation events and time patterns Tδ'(1,2,3...).The manifested
synaptic Td' structure can be considered also as a bootstrap
sequence that was generated by continuous learning processes and
perception experiences, and which, for example, serves as a
reference pattern for the event "pinprick". If a newly acquired Td
bootstrap sequence – which is given by the current properties of the
vesicle filling, as well as other significant time dependent
parameters - approximately keeps step with this existing Tδ'
(bootstrap sequence (or with a part of it), then "covariance" is
acknowledged in the synaptic structure. This opens a vesicle
attachment site at the synaptic lattice and results in the release
of all transmitter molecules that are contained in a vesicle,
whereupon an EPSP is generated at the sub-synaptic membrane
25. The potential of an EPSP corresponds to the probability
density parameters shown in Fig. 3f, which are significant for the
currently evaluated covariance. If such "probability density
parameters" sum within a certain time interval to a certain
threshold potential 27, an AP 26
is produced. This AP serves as confirmation of the event "pin
recognized", and produces a muscle reflex.
The comparison of
the current elapse time pattern with prior recorded elapse time
patterns, as shown in Fig. 3c, takes place continuously in the
synapses. Each recognized covariance of a new time sequence, that is
recorded by "temporal auto-adaptation", sets a type of "servoloop
mechanism" in motion. It initiates a process that we term "motoric
auto-adaptation", and which can be understood as the actual "motor"
in biological chemical organisms, or life forms, respectively.
Structures of temporal and motoric auto-adaptation, which are based
on STQ quantization, exist also at the lowest molecular level.
Without elapse time-supported servoloops, co-ordinated change
in biological systems would be
impossible. This applies
especially to themotion of proteins; to the recognition and
replication of the genetic code; and to other basic life processes.
The creation of higher biological/chemical order and complex systems
such as synapses or neurons presupposes the existence of an STQ
quantization molecular sub-structure, from which simple
acknowledgement and self-organization processes at a lower level
derive. Indeed, there are innumerable hierarchies of auto-adaptive
phenomena on various levels. Simple phenomena on a molecular level
also include: fusion of receptor molecules; the formation of pores,
ion canals and sub-axonal transportation structures (microtubules);
and the formation of new synapses and axonal branchings.
By this token, recognition of stimulation signal sequences by
synaptic time pattern comparison (as an involuntary reflex or as a
conscious perception), as discussed in the description of Figs.
4a - c, is an STQ-epiphenomenon. Each such auto-adaptive
STQ-epiphenomenon, for its part, is superimposed from
STQ-epiphenomena of higher rankings; for example, the analysis of
complex "time landscapes" in order to find isomorphism.
STQ-epiphenoma such as regulation of blood circulation, body
temperature, respiration, the metabolism, seeing, hearing, speaking,
smell, the co-ordination of motion, and so on, are for their parts
superimposed from STQ-scenarios of higher complexity, including
consciousness, thought, free will, conscious action, as well as an
organism's sensation of time. In all these cases, the central
nervous system looks after convergent time patterns that are placed
like pieces of a jigsaw puzzle into an integrated total sensory
scenario.
If, in any hierarchy, within a certain "latency time" (i.e. time
limit) and despite intensive "searching", no time subpattern
covariant with the STQ time pattern can be found, then the organism
displays chaotic behaviour. This behaviour restricts itself to that
synaptic part in which the non-convergence has appeared. As soon as
a covariant time pattern is found, the co-ordinated process of
temporal and motoric auto-adaptation (and auto-emulation) resumes.
(This can be likened to servo-steering that has collapsed for a
short time.) However, the "chaotic behaviour" is itself quantized as
an STQ time pattern, and is recorded by the affected synapses in
such a manner that no neurotransmitter substance release occurs
despite arriving AP's. Via subaxonal transportation structures (i.e.
the microtubules) such information streams back borne on transmitter
molecules which travel in the inverse direction along the axon.
Microtubules are used to generate new synapses and synaptic
connections at the neurons and neural networks in which a collapse
of an auto-adaptation process has occurred. The production of new
synapses proceeds to the generation of dendrites; i.e., axonal
branches that carry processing information from neurons. In this way
the auto-adaptive neural feedback mechanism regenerates itself, and
the STQ time pattern that was acquired during the short termed
"chaotic behaviour" becomes a new reference basis for the
recognition of future events. Thus, the CNS learns to record new
events and experiences; and learns to evaluate time patterns which
were unknown previously.
Fig. 5 shows a configuration in which the described invented method is applied to generate an autonomous self-organizing mechanism, in particular a robot, in which the STQ quanta are acquired by means of mechanistic sensor technology and electronic circuits. In contrast to Figs. 4a - f, in the particular case shown here, nearly exclusive STQ(i) elapse times together with STQ(v) elapse times (which are required for the measurement of the relative instantaneous speed vm) are quantized. The time data streams, designated as Tω, are obtained from these vm-adaptive STQ(i) elapse time measurements. It would nevertheless be advantageous to acquire also STQ(d) quanta, which can serve to verify the recorded time data stream Tω.
In contrast to molecular/biological organisms, in mechanistic systems it is not possible to place a comparably large number of sensors adjacent to one other on narrow sites. It is therefore necessary to acquire as many STQ elapse times as possible from the available mechanistic sensor technology, in order to attain a sufficiently large reference base for the subsequent statistical analysis. It is also worth reiterating that, as described in Fig. 3a, in multiple STQ(i) quantization, parallel and simultaneous time data are produced, so that this data must also be processed in a parallel manner.
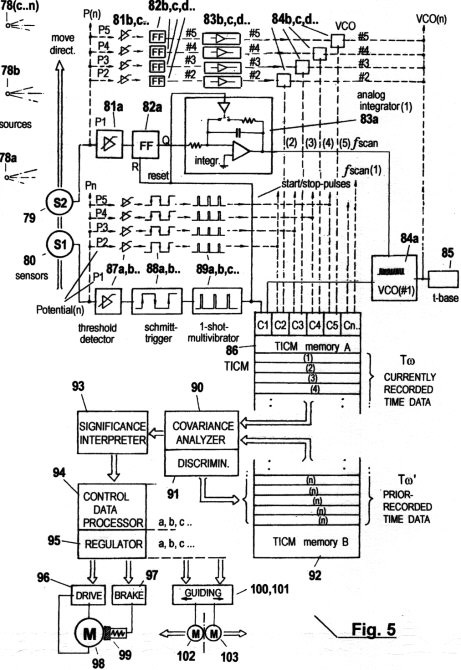
This figure shows a block diagram for a mobile autonomous robot
that has the ability to reproduce motion sequences in an
auto-adaptive manner, and to optimize the timing of its own motion
sequencesby continuous scanning and recognition of the physical
surroundings. The robotic system is equipped with equivalent
adjacent sensors 79 and 80, which
produce analog output signals, and that are inter-connected with
threshold detectors 81a,b,c,d,e... and
87a,b,c,d,e... . When sensor 79 (the
"V-sensor") moves along the corresponding external signal source
78a in the designated direction, its signal
amplitude first breaks through the lowest potential P1, which is
determined by the threshold detector 81a (see
description of Fig. 2b). The Flip-flop IC 82a
(output set to = H ) is thereby triggered. (A Schmitt-trigger IC and
a monoflop IC should be preadded in order to generate short pulses
at each phase transition.) The subsequent resettable precision
integrator IC (1) 83a provides a continually
ascending analog output signal which modulates the output frequency
ƒ of the programmable oscillator IC (VCO) The frequency ƒ is
communicated to the input of a digital TICM (a multiple time
counting andstoring IC 86 (C1)) and whereby the
current vm-adaptive time counting frequency ƒscan(1) (see also Figs.
3b,c) is produced. The integrator IC (1) 83a
therefore carries out the STQ(v) quantization. It acquires the
elapse time Tv(1) in the form of a potential increase, which is then
converted by theVCO(1) 84a into a time counting
frequency ƒscan(1), and which is inversely proportional to the
relative velocities vm(n...) with which the robotic system is moving
relative to the spatial surroundings.
After the neighbouring
sensor 80 (the "W-sensor") extends to the
perception field of the signal source 78a, its
signal amplitude first breaks through the lowest potential P1, which
is determined by the threshold detector 81a (see
description of Fig. 2b). As a result, the rising edge of the
subsequent Schmitt-Trigger IC 88a produces an
impulse in the subsequent IC 89a, whereby the
STQ(i) quantization of the vm-modulated elapse time Tw(1) is
commenced in the TICM 86(C1). Because a reset pulse
simultaneously goes to the Flip Flop 82a, causing
the analog level of the analog output of the integrator(1)
83a to be held fixed, the pulse frequency ƒ(1) persists as
a momentary vm-dependent time counting base ƒscan (1) at the output
of TICM 86(C1), and remains unchanged until the
next STQ(v)-parameter is quantized. This quantization happens
whenever the signal amplitude of the sensor 79
dropsbelow the potential P1, which is determined by the threshold
detector 81a (whence the flip flop IC 82a
is triggered by the falling signal edge), or when the sensor
79 expands into the perception field of another signal
source 78b,c,d,e...
Simultaneously an
impulse is again produced by IC's 87a, 88a and
89a, which stops the measurement ofthe elapse time
Tw (1) in the TICM 86(C1), and stores the counted
vm-modulated time pulses into the time data memory (C1). In the
memory area C1 are stored the Tw time data that refer to the lowest<
potential P1; e.g. Tw(1), Tw(8), Tw(15) etc. Quantization of all STQ
elapse times that refer to the higher potentials P2, P3, P4, P5 etc.
is handled in the same manner as for P1. When the signal amplitude
from sensor 79 passes through the threshold
potentials P2, P3, P4, P5.... (determined by detectors IC's
81b, c, d e...), the outputs of flip flops
82b,c,d,e... are sequentially triggered to = H and
therefore the subsequent integrator IC's 83b,c,d,e...
generate continuously rising analog output levels, which serve to
steadily decrease the frequencies ƒscan (produced by the VCO's
84b,c,d,e ..) until the signal amplitudesfrom
sensor 80 goes through the higher threshold
potentials P2, P3, P4, P5..(determined by detector IC's
87b,c,d,e...), when sensor 80 expands to
the perception area of the signal source 78a.
As a result, the Schmitt trigger IC's 88b,c,d,e...
are affected, and the mono flop IC's 89b,c,d,e...
produce impulses that start the acquisition of vm-adaptive elapse
time data Tw( 1, 2, 3, 4...n) in the TICM 86
(C2,C2,C3, ...Cn). The recording of these data is carried out while
the momentary vm-adaptive time counting frequencies ƒscan(1,2,3,4,.
..n) are valid, because simultaneously transmitted reset impulses to
the flip flop IC's 82b,c,d,e... hold the output
levels at the integrator IC's 83b,c,d,e... fixed,
whereby thecurrent output frequencies ƒ(1,2,3,4 ...n) are programmed
at the VCO's 84b,c,d,e... In the same mannerthe
consecutive quantization of further elapse times T( takes place when
the sensors 79, 80 move along subsequent signal
sources 78b,c,d,e... All quantized STQ(i) time date
are filed in the TICM 86(C....n). In the memory area C2 (see the
corresponding Fig. 2b) are filed the elapse times Tw(2), Tw(7),
Tw(14).. that refer to the perception area (potential) P2; in the
memory area C3 are filed the elapse times Tw(3), Tw(6), Tw(13)...
that refer to the next higher potential P3; in the memory area C4
are filed the elapse times Tw(4), Tw(5), Tw(12)... that refer to the
next higher potential P4...; and so on. The Tw-sequences currently
streaming into the TICM are generated by the current motion of the
sensor-coupled autonomous mechanism (e.g. "robot vehicle") along
some track. In the case shown, the positions of the sensors are
temporally deviating according to the positions of the external
signal sources (physical surroundings).
In the case of
absolute physical invariance between the mobile robot system and the
surroundings (so-called synchronism), no STQ parameter and no
Tw-sequence can be acquired. If such physical invariance is not
occurring, then it is possible for the autonomous vehicle to
recognize its own motion along the track by continuous comparison of
currently acquired STQ elapse time patterns Tw(1,2,3,4...n) with
prior recorded STQ elapse time patterns Tw'(nnnnn); and it is also
possible for it to perfect the recognized motions continually in an
auto-adaptive manner. A prerequisite for this is that the vehicle is
equipped with a drive and brake system controlled by data which are
calculated on the basis of continuous statistical time data
analyses.
(Compare also Figs. 3d and 3e): As soon as the
regression curve of a currently recorded time data sequence
Tw(1,2,3...) in the TICM 86 converges to the
regression curve of a previously recorded timedata sequence
Tw'(nnnn) that was acquired through a prior similar motion on the
same track, the drivesystem 98 (as well as the
brake system 99) is actuated by impulses
96, 97, which induce the autonomousvehicle to perform its
motion courses along the external signal sources
78a,b,c,d,e... in a manner such that the current motion
course is temporally and spatially approximately isomorphic to that
former motion course from which the referential time data sequence
Tw'(nnnn..) is derived. For this purpose,the TICM 86,
in which the current time data are recorded, and the memory
92, in which the prior recorded time data Tw'(nnnn..) are
stored, are interconnected with a covariance analyser 90
and discriminator logic 91, which verifies the
elapse time data and tests them for plausibility. Invalid time data
are deleted and/or interpolated, whereby no breakdown of a
data-supported servoloop can occur. Analyzer 90 and
discriminator 91 continuously scan the memory
92 with very high frequency to find approximately
covariant time data patterns. Significant data sequences are
transferred to the interpreter> that decides the respective
probability density and the value of covariance. If significant
covariance exists, then the processor 94 calculates
the appropriate actuating data for keeping an isomorphic course of
motion. These data reach the control module 95,
where they are transformed into impulses 96, 97 for
the drive and brake system 98, 99.
It is
advantageous to extend this arrangement by incorporating energetic
impulses for a steering and contra-steering system 100,101,
102, 103 that are based on the same functional principles
as above, and that are required to keep to the spatial motion course
determined by the same Tw time patterns as above. A prerequisite for
perfect functioning of such an arrangement is the utilisation of
extremely fast processors for the operation of the subsystems
90, 91, 93, 94, and 95. The
current motion course of the autonomous vehicle can be made
approximately isomorphic to the referential motion course only if
the recognition of the significant Tw '(nnnn) sequences (i.e. the
reference data), the recording and analysis of the current Tw
sequences (actual data), the computation of the control parameters
and the application of the energy impulses 96, 97
all occur nearly in real time. The vehicle would then display
behaviour similar to a "power servoloop" of the known type. This
similarity can be confirmed simply by increasingor decreasing the
base frequency fn of the clock 85, whereby the
entire temporal course in all motionphases is accelerated or
decelerated, in an absolutely synchronous manner.
Each external intervention that tries to alter or disturb the
motion course is counteracted automaticallyby the drive mechanism of
the autonomous vehicle. Therefore, an autonomous mechanism working
along these principles is comparable with a "live organism". Since
in the system components 90, 91, 93,94 and
95 a tendency is programmed that continuously optimizes the
analysis and interpretation of acquired time parameters (for
example, to allow only "authentic data"; i.e. those Tw'(nnnn) time
data that pertain to the shortest and most efficient path to
follow). In such a mechanism, there would then exist the tendency
not only for temporal and motoric auto-adaptation, but also for
optimization. (This is inherent in molecular/ biological structures
of organisms (see description to Figs. 4a - f).) The system is also
capable of determining priorities, as well as of deciding in favour
of Tw time data sequences that correspond to some other regression
curve, if an irregular track deviation that cannot be stabilized by
the control module 95 is recognized; whereupon, for
example, the vehicle emulates a new motion course and a new speed
time curve (timing). The memory of the TICM 86 can
store any alternative motion scenario in the form of Tw time data
patterns, which are accessed if a certain course deviation makes it
necessary to do so. In this way, crash situations are recognized as
soon as the danger becomes apparent, and can be avoided, since the
vehicle is ready to react in an autonomous manner.
The system goes out of control ("chaotic condition") only when
no segmental regression curve derived from prior recorded Tw-sequences
can been found that converges to a segmental regression curve derived
from currently recorded Tw-sequences. The author terms this process
"motoric auto-adaptation", or "auto-emulation". In order to be able
to identify temporal-spatial deviations of the physical surroundings
from the subjective view of the autonomous system, it doesn't suffice
in most cases just to scan external structures, land marks and light
conditions by means of optical or photoelectric sensors passively. It
is usually necessary to sense also height deviations by means of
inclination sensors; uneven surfaces by means of pressure detectors
or acceleration sensors; stationary acoustic sources by means of
microphones; gradients by means of magnet field sensors; and positions
by means of GPS; in order to acquire sufficient STQ parameters for a
reference base.
All recorded
Tw'(nnnn..) time data streams are stored in the memory of the TICM.
One can conclude from this that the adaptability and
self-organisation capability of an organism (or autonomous
auto-adaptable mechanism) increases in proportion to the quantity of
all available sensors, or, respectively, to the number of STQ
parameters that are available for the auto-adaptation process.
Another important point is that in an autonomous system, there can
be no timing without an accompanying time recording (=STQ
quantization). Auto-adaptive processes and mechanisms of the
described type will be indispensable for many future tasks in the
high technology sector; for example, in the development of
autonomous robot systems.
An example of such a task is the
following. An automobile that must find its way through traffic
autonomously, safely and efficiently, must be capable of holding
lateral and frontal distance margins, as well as speed courses,
fixed. This automobile, moreover, would have to be able to execute
autonomous overtaking procedures, and to recognize dangerous
situations in advance and avoid them. This is only possible if the
onboard computer of the vehicle is interconnected with a
multiplicity of different sensors that record a diverse variety of
signal sources; and if the vehicle is equipped with extremely fast
and efficient hardware and software that can process the STQ time
data required for auto-adaptation, approximately in real time.
Future types of microprocessors could be enhanced with hardware
structures that perform the functions described above..
Fig. 6a shows a configuration of a simple embodiment of
an aspect of the invention, in which the STQ(v), STQ(i), and STQ(d)
quantization methods introduced in Figs. 2a - c are applied to the
recognition of spatial profiles or structures. In the application
shown here, a robot arm, on which two adjacent metal sensors 104,
105 are installed at a distance b apart, must be capable of
distinguishing theprofile of the metal rail 106 while moving at
various speeds along any of the rails 106, 107, 108.
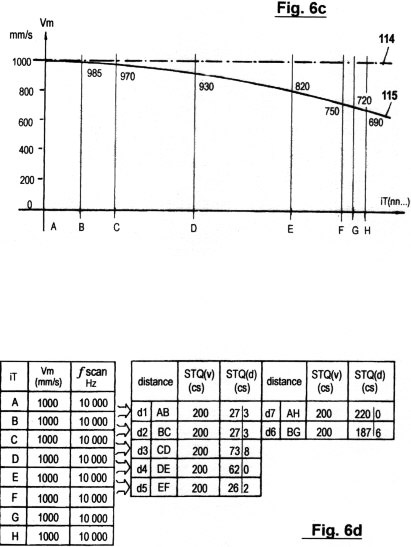
If the sensor head is moving at height h in the designated
direction, then the v sensor 104 (S2), and then the
W-sensor 105 (S1) in turn, approach the low
sensitivity area designated here as perception intensity zone
1. The lowest threshold value P1 is passed through
by the signal amplitude, and the acquisition logic 109
- mainly consisting of elements 81, 82, 83, 84, 85, 86, 87,
88, and 89 (shown in Fig. 5) - begins to
acquire v- modulated STQ(i), STQ(d) time sequences Tw(1,2,3...n) and
Td(1,2,3 ...n), which are stored in the TICM memory (A) 110.
The same time data acquisition process recurs when sensors
104, 105 meet the next higher perception area zones
2 and 3, and when the signal amplitudes
break through the potentials P2 and P3, which are preset in the
threshold value detectors.
Within the analyzer 112,
in order to identify the metal rails 106 unequivocally (which would
thereby show the characteristic profile), Tw and Td time data
streams flowing into the memory 110 must be
continually compared with the particular significant Tw', Td' time
data pattern (B) 111 that has been preprogrammed as
a "reference" pattern. Invalid or irregular time data are
recognized, then deleted or corrected by the discriminator unit
113. This unit is programmed with the capability of
improving the allocation and processing of data automatically (e. g.
verifying and checking the time data in an auto-adaptive manner) as
was already described with reference to Fig. 5. If a profile has
been "recognized", then the analyzer 112 transmits
a confirmation signal to an actuator unit of the robot, which sets a
mechanism in motion that lifts the identified metal rail up from the
ground, puts it on a conveyor belt, and so on.
Figs. 6b - e show various diagrams and charts pertaining to Fig. 6a.
Fig. 6b shows a sensometric diagram of the scanned rail profile 106. The measurement of its dimensions d1...d7 is effected exclusively utilizing STQ quanta, i.e. within the time domain. Three sensitivity zones P1, P2 and P3 are preset (in the threshold detectors as well) for profile identification. At the phase transitions (iT)A, (iT)B, (iT)C, (iT)D, (iT)E, (iT)F, (iT)G and (iT)H, digital precision timers are activated or stopped. Since the variable time counting frequency ƒscan with which these timers are counting is automatically adapted (modulated) by the current scanning velocity vm (see also Figs. 3a - g and Fig. 5), the actual dimensions d1...d7 correlate significantly with the Tw, Td elapse times that are already stored in the memory 110. As seen from the diagram, the distances AB-(d1) and BC-(d2) are obtained from STQ(d) elapse times; and the distances CD-(d3), DE-(d4), EF-(d5), as well as BG-(d6) and AH-(d7), are obtained from STQ(i) elapse times. It is to be emphasized once again that all of the (iT)n... are volatile phase transitions, and never "time points" in the classic physical understanding.
Fig. 6c shows vm diagrams of two motion courses of the sensors S1 and S2 along the metal profile being scanned. In the first case, the robot arm on which the two sensors are installed moves with an invariant speed of 1000mm/s over the profile (dash dot graph 114). In the other case, the arm decelerates from a speed of 1000mm/s at the first phase transition A to 690mm/s at the last phase transition H. The deceleration is not linear, and is shown in the graph 115.
Fig. 6d shows a fictitious frequency and time data table for Fig. 6c, with a constant vm relative speed of 1000m/s at all phase passageways (iT) A...H. Consequently, the vm-modulated time counting frequency ƒscan is 10 kHz during the entire scanning process. Because, in the case shown here, the recording ofSTQ(v) elapse time takes place with a fixed clock timing base of 200cs/b, the scanning process leads to vm-adapted STQ(d) sequences of 273cs, 738cs, 620cs and 262cs for distances AB, BC, CD, DE and EF and to vm-adapted STQ(i) sequences of 1876cs and 2200cs for the distances BG and AH. The current Tw -T d sequence, consisting of vm-adapted STQ(d) and STQ(i) elapse times, is compared in the analyzer 112 with the referential stored Tw` - Td` sequence 270, 270, 740, 620, 260, 1880, 2200, which serves as the significant time pattern, for this metal profile, that is already stored in the memory 111. If the analyzer decides that "covariance" is occurring, then a confirmation signal is transmitted to an actuator unit. The analyzer consists of comparators and/or "fuzzy logic"-IC's which ignore scattering in the boundary values (for example, decimal places are rounded up). Apart from these correction measures, tolerances, plausibility criteria and allocation criteria can also be programmed by software.
Fig. 6e shows the same frequency and time data chart as Fig. 6d, but with variable scan speed course (vm). The relative velocity of 1000mm/s at phase transition (iT)A decreases to 690mm/s at the last phase transition (iT)H. The vm deceleration is not linear. In accordance with the graph 115, at the phase transitions (iT) A,B,C,D,E,F,G,H, the momentary speeds (vm1,2,3...) are measured to be 1000, 985, 970, 930, 820, 750, 720 and 690mm/s. The vm-adaptive modulation of the time counting frequency ƒscan(1,2,3...), described above, produces phase transition values of 10, 9.85, 9.70, 9.30, 8.20, 7.50, 7.20 and 6.90kHz, which are then used to quantize the STQ(i)- and STQ(d) elapse times. Since the STQ(v) quantizations also take place with the clock time base 200cs/b, the same Tw-Td elapse time sequence for the distances AB, BC, CD, DE, EF, BG and AH results, as seen in the chart of Fig. 6d. It is obvious from this chart that the recognition of the metal profile is guaranteed, whether the vm speed course is linear or not.
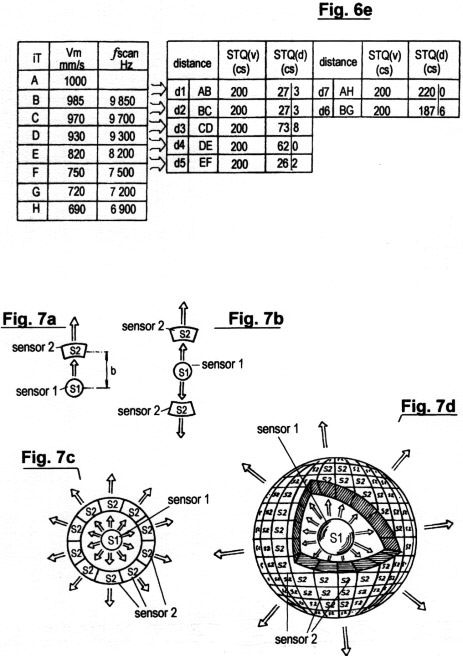
Fig. 7a - d show various configurations of sensors used
in the quantization of STQ(v) elapse times, or for the recording of
the relative speed parameters (vm), respectively. The first three
configurations show sensor constellations for 2-dimensional records
of external events. Fig.7d shows a special configuration applicable
for random 3-dimensional records of the physical surroundings.
Fig. 7a shows a sensor constellation in which a bearing, carrying
the sensors S1 and S2 on the same axis at a distance b apart, moves itself
in the designated direction along an arbitrary track; or rotates itself
about a point in space that is equidistant from both S1 (V-sensor) and S2
(W-sensor). This sensor system has only one degree of freedom.
Fig. 7b shows a sensor constellation in which a supporting
surface, carrying on the same axis two V-sensors S2 and one W-sensor S1
equidistant from each other as shown, moves itself arbitrarily in either
of the two opposite directions shown along some arbitrary track; or rotates
itself about a point in space that is equidistant from the v-sensors S2.
The sensor constellations shown in Figs. 7a and 7b are sufficient for most
robotic applications in traffic technology.
Fig. 7c shows a configuration with a number of equivalent
v-sensors S2 arranged as segments around a central w-sensor S1 on a
circular supporting surface having radius b. In this constellation,
the supporting surface can move itself in any direction in the plane
on an arbitrary track; or can rotate itself about a point in space that
is at any distance from the sensors. This sensor configuration therefore
has 2 degrees of freedom..
Fig. 7d shows a sensor configuration with a number of v-sensors
S2 arranged as segments on spherical supporting surface, with radius b,
around a central w-sensor S1. The sensor constellation can move itself
to any arbitrary position in 3-dimensional space, or can rotate in each
direction around a solid spatial point A at arbitrary distance from the
sensors. This configuration has 3 degrees of freedom.The sensor
constellations shown in Figs. 7c and 7d come into consideration primarily
for autonomous reconnaissance robots or flight objects, wherein energetic
impulses could be applied in an arbitrary direction (e.g. by means of
auxiliary rockets)..
Figs. 8a - f illustrate the configuration and functioning
principles of a further embodiment of theinvention presented herein, in
which the STQ quantization methods described in Figs. 2a, b ,c are used
to create an autonomous auto-adaptive self-organising training robot for
use in sports; a so-called "electronic hare". This system has autonomous
brake, drive and steering mechanisms, and an analyzer that continuously
compares the currently recorded vm-adaptive STQ(i)- and STQ(d) time data
patterns Tw and Td(1,2,3...) with previously recorded vm-adaptive STQ(i)-
and STQ(d) time data patterns Tw' and Td'(1,2,3....), respectively, which
serve as reference patterns. It is thereby capable of reproducing and
optimizing a motion course that has been pre-trained by the user; of
automatically finding ideal routes and speeds; of keeping distances
and times; of recognizing and warning of dangerous situations; and
of representing its own motion, as well as information about speed,
lap times, intermediate times, start to finish times, and so on, on a
monitor. It is, moreover, capable of outputting these data in an optical
or acoustic manner.
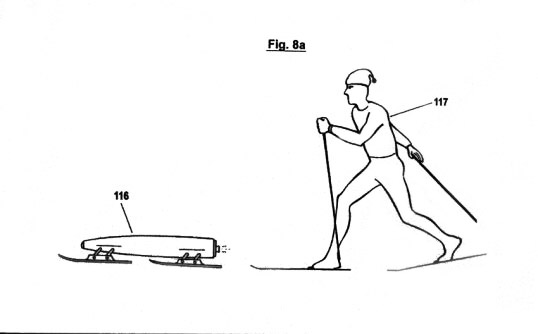
Fig. 8a shows a training robot 116 in front of a long distance skier 117. The robot vehicle envisaged for this application would be fitted with a ski undercarriage, allowing it to move with ease along snow-covered ground. It must be reasonably manoeuvrable in order to be able to match a human skier travelling in a long loop. The robot must be also able to create a new track on the same route where the former one has been covered by snow, and is therefore no longer visible. The training robot is especially suitable as an aid for blind skiers. The autonomous vehicle recognizes skiing circumstances for the blind skier, speaking out aloud hints, reports, warnings and so on by means of speech synthesis, which frees the skier and allows them more enjoyment. The robot vehicle 116 has a large number of sensors and electronic components, in the manner introduced in Fig. 5. It performs the same motion emulation, auto-adaptation and auto-optimization, often carrying out several practical tasks simultaneously. It acquires vm-adapted STQ(i)- and STQ(d) elapse time patterns from a multiplicity of sensors, compares these patterns with corresponding reference time patterns, selects the significant time data, and analyses and calculates parameters for the discrete energy impulses that manipulate the drive, brake and steering mechanisms. In the following, the essential components of the system, comprised of any of three specific types of sensors (optical, magnet field or GPS-positioning sensors) are described.
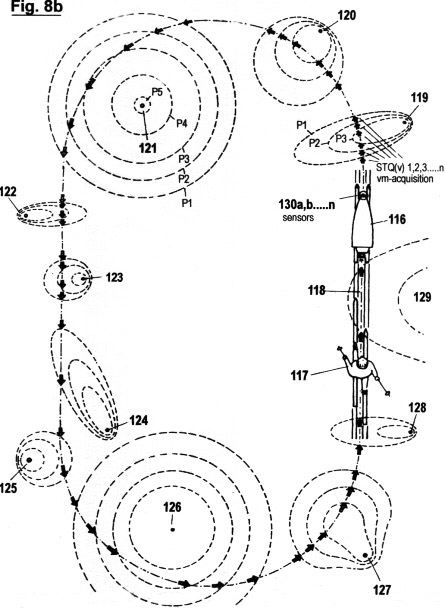
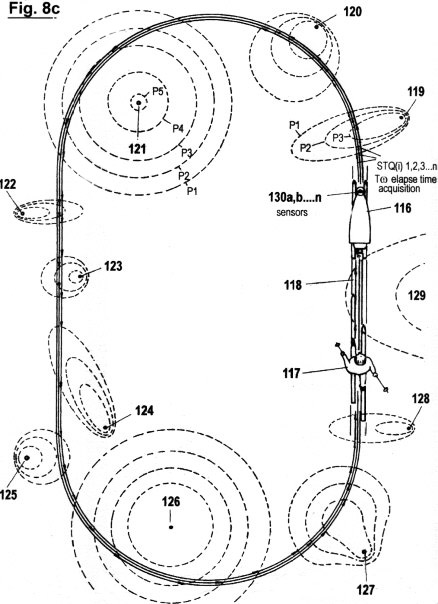
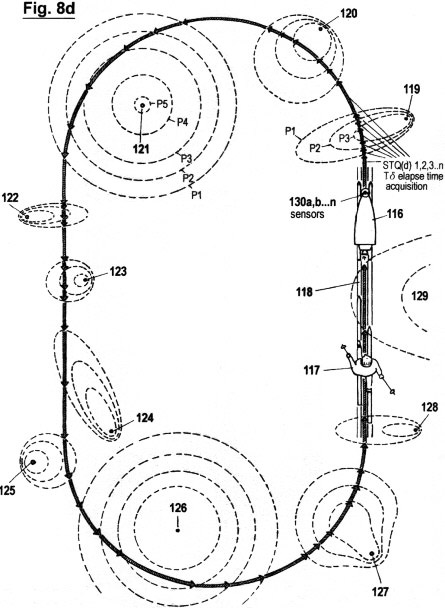
Figs. 8b-d illustrate the recording of STQ(v), STQ(i) and STQ(d) elapse times (pertaining to Fig. 8a) with use of optical or acoustic sensors. The fundamental principles of its function have already been detailed in the description of Figs. 2a - c and Fig. 5. In the present figures, the training robot (the "electronic hare") 116 is moving with variable speed in front of a long-distance skier 117 in the loipe 118. Optical or acoustic signal sources 119, 120, 121, 122, 123, 124, 125, 126, 127, 128 and 129 have been placed along the track in some arbitrary configuration, which are perceived by the corresponding sensors 130a, b,...n. At each phase transition through the threshold zones P1, P2, P3, P4, P5 etc., the designated STQ(v)- , STQ(i)- and STQ(d) elapse times are recorded. They generate the current vm-adaptive Tw '-T d '(1,2,...n) time data pattern, which is stored in the TICM. It is not crucial that the signal sources be fixed (e.g. they may be spotlights that illuminate the track for evening events). Signal sources can also be produced through differences in light intensity, contrast or colour, occurring beside trees, masts, buildings, slopes or significant land marks in daylight. Headlights could even be installed on the training robot itself, whereby the optosensoric recording of the reflected light and the evaluation of the light structures of the spatial surroundings may be used for recognizing its own motion. The same set-up may be used also with ultrasound sensors. On the other hand, acoustic signal sources could equally well be of natural origin; for example, the sounds of a brook running beside the loipe, or a waterfall. Generally, any volatile combination of light and shadow, or any noise source, can be decisive in the recognition of a certain object. The particular identity of the object may be determined by comparison of vm-adaptively recorded STQ(i)- and STQ(d) elapse time patterns with the Tw '-T d '(1,2,3...n) patterns, which are stored in the TICM and which represent each individual external object. In order to simplify the present description and demonstration, it is assumed that the signal sources 119 ...129 in Fig. 8b are lamps installed along the robot's route, making it possible for the robot to use the loipe at twilight or in darkness. According to the primary domain of application of such a robot, the training robot 116 skis with precision behind the skier 117 along the skier's track, with all STQ time data vm-adaptively recorded and stored in the TICM working memory (see also Fig. 5). The distance between robot and user is precisely controlled by a distance sensor. However, in order to be able to invoke the robot vehicle's drive, brake and steering mechanism, STQ time data that could serve as reference data must already have been loaded into the TICM prior to the journey. Therefore, as a first step, the acquired time data are stored in the TICM reference memory; i.e., Tw -T d (1,2,3...) are mapped to Tw '-T d '(1,2,3...) initially. Subsequently, the emulation of the skier is repeated several times, with increasing processing speed as the robot learns more about the skier, and with variable speed and track courses; whereupon more and more covariant Tw '-T d ' time data patterns are contained in the reference data memory, which the robot's discriminator and analyser can access (see also Fig. 5). The interpretation and optimization program is put into action, which filters through only "authentic" T w '-T d ' time data that are deemed to pertain to the best and most efficient trajectory of motion, and which eliminates at the same time those data recognized as "irrelevant". This resembles a "learning process" that the robot vehicle has to undertake until it can finally ski "autonomously"; i.e. relatively freely, and in accordance with self-appropriated patterns and self-decided criterions, without any remote control or regulation by a pre-programmed algorithm. Upon reaching this stage, the training robot functions as a "trainer" or "pilot" who has the task of helping the user find ideal speeds, the best track and optimal timing. This optimal information that is communicated to the user is only that which has been learned by the robot itself. The training robot continues to improve itself also during this "practical work" (i.e. while helping the user), in continually optimising and supplementing the STQ reference data stored in the TICM. The ability to identify and recognize trajectories of motion or external signal courses and objects is always upgradeable. It depends on the quantity and variety of sensors used, as well as on the memory capacity of the TICM. Thus it is possible to induce the robot vehicle to recognize dangerous situations and to warn the user acoustically or optically; and to keep distances and times more exactly. In the present application, the vehicle performs automatic tracking and motion emulation along a loipe, even if the original track has been covered by snow and is no longer visible. Additionally, the robot vehicle has a monitor on which its own motion relative to its spatial surroundings can be visualised; as well as electronic measures to output speeds, lap times, intermediate times, total times or other relevant data in an optical or acoustic manner. An essential property of the robot vehicle shown here is that a simple adjustment (increase or decrease) of the central clock frequency can synchronously accelerate or decelerate the entire temporal course of all motion components (see also Fig. 5). For instance, this property is necessary in order to adapt the speed of the training robot in all sections according to the physical fitness of the user. This can happen manually by a remote control device, or automatically; for example, by a frequency or blood pressure data transponder.
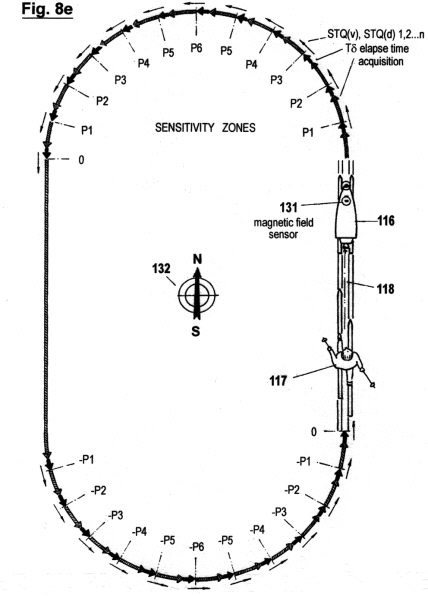
Fig. 8e shows the recording of STQ(v) and STQ(d) elapse times for the robot in Fig. 8a in the case when magnetic field sensors are installed. The signal source here is assumed to be the earth's magnetic field. In the example shown here, where the track forms a closed loop, the quantization of STQ(i) elapse times is inefficient, and therefore not undertaken. In the illustrated picture, the training robot ("hare") 116 is moving autonomously with variable speed in front of the long distance skier 117 along the loipe 118. Various vehicle position readings are produced along the track, with variable gradients to the earth's magnetic field 132. The magnitude of these gradients are acquired by the magnet field sensor 131. In this particular example, the magnitude follows a sinusoidal course. At each phase transition to the threshold zones P1, P2, P3, P4, P5, P6, and so on, the STQ(v) and STQ(d) elapse times are vm-adaptively recorded, which provides the current T( time data pattern that is stored in the TICM. The additional quantization of STQ elapse times from magnetic field gradients helps to locate covariant T w '-T d ' time patterns that are stored in the reference data memory. Consequently, the auto-adaptation and recognition capability of the robot vehicle is improved. The more sensors involved in the auto-adaptation process, the more "autonomous" is the described mechanism (see also Fig. 5). A self-organizing, autonomous organism based on biological or chemical structures, as discussed in Figs. 4a -f , can be produced in this manner.
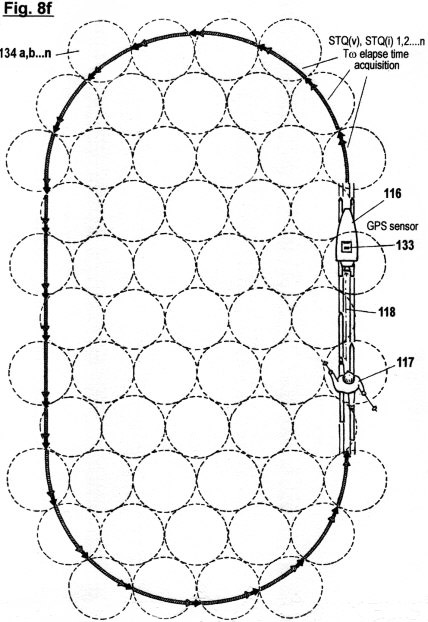
Fig. 8f shows the acquisition of circular position fields by means of GPS sensors. These measurements (in addition to those shown in Figs. 8b - e) are used to improve temporal and motoric auto-adaptation and make auto-covariance behaviour and motion emulation more precise. A prerequisite for successful function is a GPS ("global positioning system") of high quality, which operates with extremely low errors. Since a square wave signal is received in this case (therefore no subdivision into distinctive sensitivity zones is possible) only STQ(v)- and STQ(i) elapse times, but no STQ(d) elapse times can be quantized - which, as we have seen, are measured between phase transitions from lower to higher potentials, and, respectively, vice versa. In Fig. 8f the training robot ("hare") 116 moves itself with variable speed in front of the long distance skier 117 along the loipe 118, while circular GPS position fields are produced along the track 134a,b,...,n, which are perceived by the GPS sensor 133 with high precision in a reproducible manner. The radii of the position fields, as well as the resolution between adjacent fields, is adjustable. With each detection of a new position field, a trigger signal is transmitted to the STQ acquisition unit, which records the STQ(v) and STQ(i) elapse times, and which then stores these currently vm-adaptive recorded time data sequences T w (1,2,3....) into the TICM. The ability of the robot to otimize auto-adaptation can be aided by counting and comparing the number of detected position fields, or by assigning a specific data code to time data within each crossed position field.
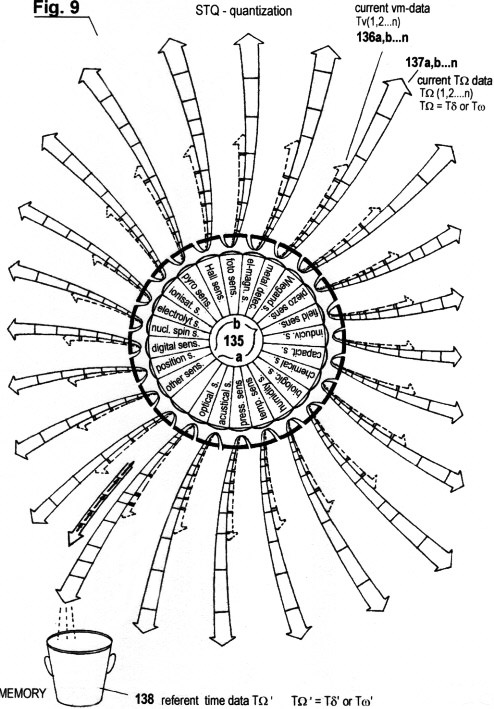
Fig. 9 is a schematic diagram showing how time data streams are produced. Each transition of the amplitude through sensitivity zones or threshold potentials in redundancy-poor autonomous self-organized systems (such as mechanistic robot systems or organisms) leads to the quantization of elapse times, if these systems are equipped with sensors (or receptors) that are adequate for the perception of the external physical surroundings. It is asserted that the core technology shown in the diagram has universal validity and applicability. The diagram shows a highly simplified scheme for the technology, which can be understood plainly by a non-expert.
The principles of this invention, as represented schematically in this diagram, are summarized below:
1) The "primary act" of every autonomous organism (including autonomous self-organizing robots) is to "explore" their surroundings in order to ascertain whether temporal-spatial variation exists between its own physical state and that of its surroundings. In order to do this, a multiplicity of sensors or receptors 135a, b...,n are necessary.
2) Only when deviation exists, are the current STQ elapse times Tw(1,2...n) or Td(1,2...n) 137a,b,...,n derived. The time counting frequency of their measurement depends on currently acquired STQ(v)- quanta Tv(1,2,3....n) 136a,b,c,.....n, which represent para- meters for the temporal-spatial variations vm(1,2 ...n) between sensors 135a,b,....n and external signal sources. These deviations are identical to the "relative speeds" vm(1,2,...n). Note: vm(1,2,...,n) are always acquired by means of an invariant time counting frequency f, respectively, at an absolute time base.
3) The current STQ elapse times Tw(1,2..n) or Td(1,2..n) flow into so-called "information pots" 138 (or time data memories) and form STQ time data patterns Tw'(1,2....n) or Td'(1,2...n), which serve as reference patterns. If the organism finds sub-sequences of these Tw' or Td' patterns which in some combination are covariant with a currently recorded Tw or Td - pattern, then the organism interprets these combinations of sub-sequences as an "iso- morphous pattern" significant for defining the "actually perceived event-pattern" (i.e. what actually is). In this way, the present event (represented by temporal or spatial deviations between sensors and external signal sources) is "recognized".
4) An organism is equipped with "actuators" that influence a self-referential change - that is concurrently being recognized - in an organism's temporal-spatial condition (e.g. its own motion) in such a manner, that the change is highly covariant with a prior recorded pattern of change of a temporal-spatial condition (it emulates the prior pattern). Because the shortest and most efficient time patterns have a tendency to be of high priority while new Tw or Td sequences are being recorded in the memory, organisms continuously try to optimize changes in temporal-spatial conditions. Both processes result exclusively from comparison of quantized STQ elapse times and from recognition of isomorphous time data patterns (see also Fig. 5), and are termed "auto-emulation" and "auto-optimization"; or, equivalently, "autocovariance behaviour".
5) An essential consequence of these considerations is that a teleological tendency inheres in generates the ability for self-organisation.
6) As seen from Fig. 10, both "time" and "velocity" unequivocally depend on the existence of sensors for their perception. Actually, all time data and information flow from the "present" (the origin of the recording) into the "past" (the verifiable existence). Indeed, time and velocity are not "sensed" as a continuum, but in the form of quanta. In order to feel both physical quantities as a continuum, an enormous capability for auto-adaptation and auto-emulation is required of an organism. It can be said that the above fundamental principles are valid not only for robotics and biological units but also for molecular atomic and subatomic structures. Also, these have to be "time sensing organisms" otherwise they can have no basis for existence. Consequently: time, space - every physical quantity – only sensorial together with distinct sensitivity zones; and these form the basis for local subjective time sensing together with a general universal tendency for auto-adaptation, auto-optimisation, and auto-emulation. This is a fundamental teleological principle.
FINAL SUMMERY
1) The herein described invented method is universally applicable and describes the ultimate achievable mechanisms and organisms.
2) Discrete time quantization methods, according to which the received signal is scanned and digitized at predetermined points in time, prove themselves to be inadequate in the generation of highly efficient autonomous self-organisation processes.
3) In redundancy-free autonomous self-organizing systems, there are no "points in time" and there is no determinism. In these systems, STQ elapse times are quantized which are derived from the temporal/spatial changes in physical conditions between sensors and external sources.
4) Each such system has its own time counting pulses and produces its own time. The time counting frequency for the quantization of elapse times is continuously adapted in an auto- adaptive manner according to the relative velocity vm with which changes in condition occur. The time recording has in each case a quantum nature; i.e. it has the properties of a "discrete counting", no matter whether the recording is analogue or digital. Moreover, the time recording is subjective and passive; i.e. the time quanta are "sensed" and not "objectively measured" as in the conventional physical understanding.
5) In order to be able to quantize elapse times in autonomous self-organising systems, the individual receptors or sensors must have distinctive grades of perception zones (or threshold values).
6) In order to explain precisely the difference between "synchronism" (in the conventional understanding) and "auto-adaptation", we define the following: a) parallel synchronism (i.e. "synchronism"): this occurs when temporal changes of physical conditions of different systems are covariant at the same time. b) autonomous adaptation (i.e. "auto-adaptation"): this occurs when temporal changes of the physical state of a particular system are covariant at different times.
7) In all redundancy-free autonomous systems the capability for self-organisation increases with the quantity of elapse time parameters available for autonomous adaptation and for optimi- zation process, as well as with the number and variety of sensors or receptors.
8) With synchronism (definition 6a above), the number of quantized elapse time parameters vanishes; in 3b this number is a maximum (and point 7 above is valid! ). Therefore one can conclude that there is an inherent tendency in all autonomous systems of the type discussed herein, towards continuous auto-adaptation, auto-optimization and auto-emulation. This is similar to the biological term "vitality".
9) In autonomous self-organizing systems, there is no "timing" (i.e. temporal motion coordina- tion) without the comparison of currently acquired elapse time patterns with previously recorded elapse time patterns. Briefly stated, there is no "timing" without accompanying "time keeping".
10) Auto-adaptation theorem of Erich Bieramperl :
Every current non-chaotic change (A) in condition of an autonomous system (X) with the variable dynamic trajectory vm(1,2,3....n) underlies a currently acquired sequence of elapse times TW(1,2,3 ...n) as well as a covariant sequence of elapse times TW'(1,2,3 ...n) from a temporal displaced condition change (A') or from a combination of distinct temporal displaced condition changes (A1 ') (A2 ')...( An'), whereupon (A) with (A') or (A) with (A1') (A2') ....(An') are approximately isomorphous. Hence: TW = vm adaptively acquired current STQ(i) or STQ(d) elapse times Tw or Td TW' = vm adaptively acquired covariant STQ(i) or STQ(d) elapse times Tw' or Td' .
Other consequences in the scientific domain are the following:
11) Each preselection of a certain time for an intended action, a so-called "act of free will" by an autonomous organism, results from continued autonomous adaptation of the described type, and is therefore not realizable in a deterministic manner.
12) From the ability of an autonomous system to find previously acquired elapse time patterns matching with currently acquired elapse time patterns, and from trying to emulate these, not only is auto-adaptation, auto-optimization, self-organisation and recognition of physical surroundings and self-motion made possible, but ultimately also motion co-ordination (timing), intelligent behaviour and conscious action are produced.
13) Auto-adaptive, auto-optimizing and self-organizing processes of the described type have universal validity not only in autonomous mechanistic systems, robots, automatic machines biological organisms, but also in molecular and atomic structures. All autonomous self- organizing systems contain information in form of time data.
The following results from the property that in such systems, "time" is "subjectively sensed" and not "objectively measured ":
14) In the universe, all time dependent physical values are "subjectively sensed". If there is no adequate sensorium for time and velocity, then "time" cannot exist objectively. Example: in "black holes", no "time" exists because there is no sensorium for it. In this case, the atomic and subatomic sensorium is quasi "dead". Each change of physical condition, which does not underly an auto-adaptive process, continues increasingly chaotically; whereupon it follows that the described tendency for auto-adaptation in the universe counteracts the tendency towards entropy and chaos.
15) If vm is too high and STQ(v) is too short to be measured (or "sensed"), then neither an auto- adaptation nor any self-organization process results (because no elapse times are derivable). Therefore, for example, the velocity c of propagation of light is an "ultimate value", because it implies the shortest STQ(v) quantum that can be "perceived" by atomic structures.
16) If there is absolute physical invariance between the sensorium of autonomous systems and their surroundings, then also no STQ quanta are derivable. This is the reason why, for example, absolute zero ( 273,15°C) is an ultimate physical quantity. In this case, the atomic and subatomic sensorium is not capable of recognizing a lower temperature because of lack of STQ quanta, and no auto-adaptation process can take place.
17) As mentioned before, atomic and subatomic structures also display sensory and time quantization properties. Their description from the view of quantum theory is inadequate. If there is no measurement or observation of an event, then exists also neither "time" nor velocity" (S.13). Quantum phenomena appearing in the known two slit experiment or in the SCULLY experiment (quantum indeterminism) are explicable in this way.
18) The electromagnetic force, gravitation, the strong and weak interaction (nuclear force), so- called "autocatalysis" (KAUFFMAN), "synergetic effects" (HAKEN), or other phenomena are produced by the existence of time quantization sensorium, auto-adaptation and auto- emulation. These features can be regarded as the inherent teleological principle of the universe (S. 8).
19) The ability to perceive time and velocity as a continuum, and not as an endless series of sensed elapse times, is likewise produced from continued auto-adaptation and self- organization processes. The higher the "intelligence" of an autonomous system as a result of such processes, the more distinctive its subjective time perception and its ability to anticipate.
Consequences for metamathematics, propositional calculus, epistemology and philosophy are:
1) Because there are no deterministic point of times, the status of a system can neither be ascertained to be at a certain "point in time", nor "points in time" can be determined for a future status. There is nowhere any type of determinism. Since the classical physics as well as the quantum theory are based on the postulate that a system is in a certain status at a certain "point in time" (in the first case as points of phase space, and in the other case as probability distributions in phase space), neither theory can be completely consistent (see also THOMAS BREUER / 1997).
2) Regarding WIGNER (1961), an absolutely universally valid theory would have to be capable
of describing the origin of human consciousness. The auto-adaptation theory described
herein could be capable of this; the quantum theory cannot. (Wigner postulated that
complex quantum mechanics delivers a usable description of the physical reality only
when there is no "subjective sensing". The author holds the view that subjective sensing
also exists in atomic and subatomic structures).
3) Sequences of elapse times like TW and TW' are definable as strings of an axiomatic formal
system; albeit this system is a "time domain system" and not an arithmetic systems in the
usual sense of the classic number theory. Indeed, said formal system shows at least one
axiom and derives from it continuous strings of numbers through the application of a certain
algorithm. Regarding TURING, an axiomatic number theoretical system can be produced
also by a mechanical procedure, which produces "formulas and algorithms".For this reason,
the known logic theorems of GOEDEL, TARSKI or HENKIN are absolutely applicable on
such a model. GOEDEL`s incompleteness theorem shows that each extensive number
theoretical model includes consistent formulations which cannot be proven with the rules
of the model, and which therefore are undecidable. This is valid also to metatheoretical
models and to meta-metatheoretical models etc.
For example, a self-referential metatheoretical sentence like the type of the Goedel formulation
"I am provable" is neither provable nor disprovable. A decision procedure for this
proposition leads to an infinite regress. TARSKI showed that a decision procedure for
number theoretical "truth" is also impossible, and leads to an infinite regress. Thus, a self-
referential sentence of the type "I am provable" is admittedly "true", but not "provable". It
follows, that "provability" is a weaker notion than "truth". HENKIN showed that there are
sentences, that assert their own provability and "producibility" in a specific number theore-
tical model and which are invariable "true". A self-referential sentence based on Henkin`s
theorem would be: "It exists a number theoretical model in which I am provable". Strings
of quantized elapse times like TW and TW' approach the domain of validity of HENKIN`s
theorem. Applying Henkins logic, these strings assert: "I will be produced to proved". TW
and TW's are therefore strings or sentences that are produced in a specific formal model,
which induces its own decision procedure on truth, consistence, completeness and
provability through continued self-generation (see also description to Fig.10).
In contrast to self-referential strings or sentences of the Gödel or Henkin type, strings of
elapse times are never asserted to be "true", "consistent", "complete" or "provable" to a
certain "point in time", because within the "number theoretical model" in which they are
produced, no "points of time" exist. This model also prohibits superior semantics or
metatheories or meta-metatheories. It is plainly obvious that each formal system, each
metatheory, each meta-metatheory and each semantics, in which axioms, strings or
sentences of any type are formulated, is the result of continued autonomous adaptation
(which is based on the quantization of elapse times) and therefore a derivation of the
model described in this work.
4) The cognition, that a specific formal system exists asserting absolute universal validity, from
which everything has been produced and to whom all other systems have to be subordinated,
is not new. Already in early antiquity, many years before PLATO and ARISTOTLE, the
Hebrew Scriptures (2. Moses 3: 14) let this "source of all logic" say from itself: "JHWH"
(spoken: Jahwe or Jehovah), that is about: "I shall be proved". This sentence asserts its own
decision procedure on provability, truth, completeness and consistence; through a specific
formal system, that it "induces to be".
5) There is no "cognition" without "recognition"
[1] Thomas BREUER (1997) “Quantenmechanik: Ein Fall für Goedel” ISBN 3-8274-0191-7
[2] Eugene WIGNER (1961) “Remarks on the Mind-Body-Question”,
siehe auch: Roger Penrose: Des Kaisers neue Kleider”/ Spektrum-Verlag Heidelberg (S. 287)
[3] Kurt Goedel “Über formal unentscheidbare Sätze der Principia Mathematica und verwandter Systeme I. (1931), siehe auch: Douglas HOFSTADTER “Goedel, Escher, Bach” ISBN 0-394-74502-7 (Seite 19)
[4] Douglas HOFSTADTER “Goedel, Escher, Bach” (s. Seite 618: “Tarski`s Satz”)
[5] Douglas HOFSTADTER “Goedel, Escher, Bach” (s. Seite 577: “Henkin-Sätze”)
[6] Siehe WIKIPEDIA unter JHWH
Please direct any questions to: info@sensortime.com